Готхаб, горад, адміністрацыйны цэнтр Грэнландыі, у Готхабскай бухце на зах. узбярэжжы вострава. Засн. ў 1727. Каля 10 тыс.ж. (1998). Марскі порт. Аэрапорт. Цэнтр рыбалоўства і рыбаперапрацоўчай прам-сці. Вытв-сць марожанага філе рыбы. Ун-т. Выд-вы, радыё- і тэлестанцыі.
злучэнні, якія складаюцца з астаткаў азоцістай асновы і вугляводу рыбозы (рыбануклеазіды) ці дэзоксірыбозы (дэзоксірыбануклеазіды); N-гліказіды пурынавых або пірамідзінавых асноў. Найб. значэнне маюць Н., што ўваходзяць у склад нуклеінавых кіслот: адэназін, гуаназін, урыдзін, цытыдзін, цымідзін. Свабодныя Н. ўтвараюцца пры гідролізе нуклеінавых к-т і нуклеатыдаў. Абарачальнае фасфат-залежнае расшчапленне Н. да рыбоза-1-фасфату і свабоднай асновы адбываецца пад уздзеяннем ферментаў нуклеазідфасфарылаз. Спецыфічныя нуклеазідазы каталізуюць гідроліз Н. і разрываюць сувязі паміж цукрам і асновай. Фасфарыліраванне Н. кіназамі выклікае ўтварэнне нуклеатыдаў. Некат. пурынавыя і пірымідзінавыя Н. з хім. мадыфікаванай вугляводнай ч. малекулы і (або) асновай валодаюць антыбіятычнай актыўнасцю і блакіруюць абмен пурынаў, пірамідзінаў і бялкоў як антыметабаліты натуральных субстратаў. Яны знаходзяцца ў клетцы ў свабодным стане і не ўваходзяць у склад нуклеінавых к-т.
складаныя комплексы нуклеінавых кіслот з бялкамі. Адрозніваюць Н. ў комплексе з ДНК і РНК. Дэзоксірыбануклеапратэіды знаходзяцца ў ядрах клетак (рэчывы храмасом), мітахондрыях і галоўках сперматазоідаў. Іх бялкі з’яўляюцца гістонамі і пратамінамі (нізкамалекулярныя бялкі), размяшчаюцца ў жалабках двайной спіралі ДНК, стабілізуюць яе структуру і рэгулююць матрычную актыўнасць; звязаны з нуклеінавай к-той электрастат. узаемадзеяннем. Рыбануклеапратэіды ўваходзяць у склад рыбасом, вірусаў, інфармасом. Кожная з гэтых структур мае адну або некалькі малекул РНК і дзесяткі розных бялкоў. Вылучаныя і ачышчаныя кампаненты рыбасом і вірусаў здольныя да самазборкі і ўтварэння біялагічна актыўных сістэм (напр., інфекц. вірусаў).
фосфарныя эфіры нуклеазідаў. Складаюцца з азоцістай асновы (звычайна пурынавай або пірымідзінавай), вугляводу рыбозы (рыбануклеатыды) ці дэзоксірыбозы (дэзоксірыбануклеатыды) і аднаго або некалькіх астаткаў фосфарнай к-ты. Уваходзяць у склад нуклеінавых кіслот, найважнейшых каферментаў і інш. біялагічна актыўных рэчываў. Нуклеазідтрыфасфаты — Н., што маюць 3 астаткі фосфарнай к-ты, з’яўляюцца макраэргічнымі злучэннямі, крыніцамі і пераносчыкамі хім. энергіі фасфатнай сувязі. Асобую ролю адыгрывае адэназінтрыфосфарная кіслата — універсальны акумулятар энергіі, які забяспечвае розныя працэсы жыццядзейнасці (сінтэз бялкоў, поліцукрыдаў, ліпідаў і інш.). Нуклеазідтрыфасфаты з’яўляюцца таксама субстратамі для сінтэзу нуклеінавых к-т. Да важнейшых рэгулятарных Н. адносяцца і іх цыклічныя формы. Свабодныя нуклеазідмонафасфаты ўтвараюцца шляхам сінтэзу або пры гідролізе нуклеінавых к-т, далейшае іх фасфарыліраванне прыводзіць да ўтварэння адпаведных нуклеазідды- і нуклеазідтрыфасфатаў. Распад Н. адбываецца пад уздзеяннем нуклеатыдаз і нуклеатыдпірафасфарылаз.
гетэралітычныя рэакцыі арган. рэчываў з нуклеафільнымі рэагентамі (нуклеафіламі). Нуклеафілы (ад лац. nucleus — ядро і грэч. phileo — люблю) — аніёны (Cl−, OH−, CN−; NO2−, OR− і інш.), а таксама неарган. і арган. малекулы, што маюць свабодную пару электронаў (напр., вада, аміяк, аміны, спірты, фасфіты). У арган. сінтэзе пашыраны нуклеафільнага далучэння рэакцыі па карбанільнай групе, ці кратнай сувязі вуглярод — вуглярод і замяшчэння рэакцыі, характэрныя пераважна для аліфатычных злучэнняў.
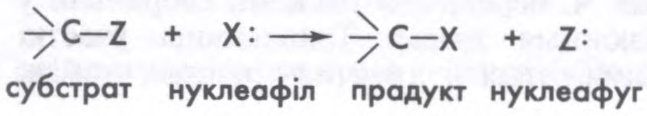
Пры рэакцыі нуклеафільнага замяшчэння ў насычанага атама вугляроду нуклеафіл (X:) аддае субстрату сваю пару электронаў на ўтварэнне сувязі C—X і пры гэтым выцясняе групу Z:, ці нуклеафуг (ад лац. nucleus + fugio збягаю), з парай электронаў, што звязвалі яе з атамам вугляроду. Скорасць і механізм рэакцыі вызначаюцца рэакцыйнай здольнасцю нуклеафілу (нуклеафільнасцю) і нуклеафугу (нуклеафугнасцю), будовай малекулы субстрату і ўмовамі рэакцыі (растваральнік, т-ра, ціск і інш.). Адрозніваюць мона- і бімалекулярны механізмы. Пры монамалекулярным (SN1) механізме спачатку (пад уздзеяннем растваральніку) утвараецца трохкаардынацыйны карбкатыён і нуклеафуг (стадыя, якая звычайна вызначае скорасць усяго працэсу), а затым нуклеафіл хутка далучаецца да карбкатыёна; атака нуклеафілу роўнамагчымая з абодвух бакоў рэакцыйнага цэнтра і ў выпадку яго асіметрыі прыводзіць да рацэмізацыі. Пры бімалекулярным (сінхронным) замяшчэнні (SN2) атака нуклеафілу ідзе з боку процілеглага адносна групы, што адыходзіць, і прыводзіць да змены абс. канфігурацыі асіметрычнага цэнтра малекулы (т.зв. вальдэнава абарачэнне). Гл. таксама Рэакцыі хімічныя.
біяпалімеры, якія маюць у сабе фосфар і універсальна распаўсюджаны ў жывой прыродзе. Адкрыты швейц. біяхімікам І.Ф.Мішэрам (1868) у клетках, багатых ядзерным матэрыялам (напр., лейкацыты, сперматазоіды ласося). Лінейныя малекулы Н.к. утвораны нуклеатыдамі, а эфірныя сувязі паміж вугляводам аднаго нуклеатыду і фасфатам другога ўтвараюць іх вугляводна-фасфатны шкілет. Высокапалімерныя ланцугі Н.к. маюць ад некалькіх дзесяткаў да сотняў млн. нуклеатыдных астаткаў, іх малекулярная маса 105—1010. Звычайна Н.к. маюць астаткі дэзоксі- або рыбануклеатыдаў у якасці манамераў. У адпаведнасці з гэтым адрозніваюць дэзоксірыбануклеінавыя кіслоты (ДНК) і рыбануклеінавыя кіслоты (РНК). Малекулы ДНК маюць 2 ланцужкі, РНК пераважна адналанцужковыя. Першасная структура Н.к. — паслядоўнасць нуклеатыдаў у неразгалінаваным полінуклеатыдным ланцужку. Спецыфічная паслядоўнасць азоцістых асноў функцыянальна значная і унікальная для кожнай Н.к., абумоўлівае вял. разнастайнасць індывід. малекул ДНК і РНК і адначасова — відавую спецыфічнасць (Н.к. кожнага віду маюць пэўны нуклеатыдны склад). Другасная структура Н.к. — прасторавае размяшчэнне нуклеатыдных звёнаў — узнікае ў выніку міжплоскасных узаемадзеянняў суседніх асноў і ў выпадку т.зв. камплементарнага спарвання вадародных сувязяў паміж процілеглымі асновамі ў паралельных ланцугах. Адрозненні ў структуры манамерных звёнаў вызначаюць розныя хім. ўласцівасці і макрамалекулярную (прасторавую) структуру абодвух тыпаў палімераў. У склад клетачных арганізмаў уваходзяць ДНК і РНК, вірусы маюць Н.к. аднаго тыпу. Біял. роля Н.к. заключаецца ў захаванні, рэалізацыі і перадачы спадчыннай інфармацыі, якая «запісана» ў паслядоўнасці нуклеатыдаў (генетычны код). Структурныя кампаненты Н.к. выконваюць функцыі каферментаў, уваходзяць у іх склад, прымаюць удзел у абмене рэчываў, акумуляванні, пераносе і трансфармацыі энергіі. Параўнальны аналіз Н.к. у розных групах арганізмаў выкарыстоўваецца ў даследаваннях па сістэматыцы і эвалюцыі (ступень падабенства ў будове Н.к. вызначае ўзровень філагенетычнай блізкасці арганізмаў). Вывучае Н.к. малекулярная генетыка і малекулярная біялогія.
Літ.:
Шабарова З.А., Богданов А.А. Химия нуклеиновых кислот и их компонентов. М., 1978;
Ленинджер А. Биохимия: Пер. с англ.М., 1976;
Уотсон Дж., Туз Дж., Курц Д. Рекомбинантные ДНК: Пер. с англ.М., 1986;
Зенгер В. Принципы структурной организации нуклеиновых кислот: Пер. с англ.М., 1987.
каменны, найчасцей крамянёвы абабіты жаўлак, ад якога ў каменным веку адбівалі або адціскалі пласціны ці адшчэпы для вырабу прылад працы. У сярэднім палеаліце былі пашыраны дыскападобныя адна- і двухбаковыя Н., якія аббівалі па акружнасці, у познім плеаліце — прызматычныя Н., у канцы палеаліту, у мезаліце і неаліце — пірамідальныя, клінападобныя і аморфныя Н. Часам сам Н. выкарыстоўвалі як ударную прыладу працы.