пэўная канфігурацыя кропкавых крыніц поля (зарадаў). Напр., кропкавы зарад — М. нулявога парадку; 2 процілеглыя па знаку і роўныя па модулі зараду — дыполь, ці М. 1-га парадку; 4 аднолькавыя па модулі зараду, размешчаныя ў вяршынях паралелаграма так, што кожная старана злучае процілеглыя па знаку зарады — квадруполь, ці М. 2-га парадку. Асн. характарыстыка М. — мультыпольны момант, які дазваляе адназначна звязаць палі М. з палямі складаных сістэм крыніц на вял. адлегласцях, што вядзе да уніфікацыі рашэння шырокага класа задач. Паняцце М. дастасавальнае для апісання палёў, створаных эл., магн. і інш. зарадамі.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДЭ́ЛЬТА-ФУ́НКЦЫЯδ-функцыя, δ-функцыя Дзірака,
сімвал, што выкарыстоўваецца ў матэм. фізіцы пры рашэнні задач, у якія ўваходзяць засяроджаныя велічыні (маса, зарад, нагрузка і інш.). Абазначаецца δ(x).
Д.-ф. можна вызначыць як шчыльнасць размеркавання масы, калі ў пункце x=0 засяроджана адзінкавая маса, а ва ўсіх астатніх пунктах маса роўная нулю. Таму δ(x) = 0 пры x≠0 і δ(x)=∞ пры x=0, прычым
. Такім чынам, Д.-ф. можна разглядаць як бясконцы ўсплёск адзінкавай інтэнсіўнасці. Асн. ўласцівасць Д.-ф., што вынікае з гэтага вызначэння:
, дзе 𝑓(x) — кожная неперарыўная функцыя.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
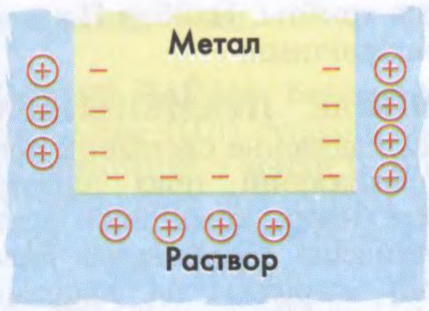
ПАДВО́ЙНЫ ЭЛЕКТРЫ́ЧНЫ СЛОЙ,
сукупнасць эл. зарадаў процілеглых знакаў, размешчаных уздоўж мяжы падзелу фаз. Утвараецца электронамі, іонамі, а таксама арыентаванымі палярнымі малекуламі, якія маюць уласны дыпольны момант. Абумоўлівае электракінетычныя з’явы.
Будова П.э.с. істотная для эл.-хім. рэакцый, для электролізу і інш.Напр., у хім. крыніцах току (гл.Гальванічны элемент) пры апусканні металу ў электраліт метал «аддае» дадатныя іоны, а сам набывае адмоўны зарад. Дадатныя іоны электраліту прыцягваюцца паверхняй металу і ўздоўж паверхні дакранання фаз утвараецца П.э.с. Эл. поле, якое ўзнікла паміж зараджанымі слаямі, перашкаджае далейшаму растварэнню металу і пры пэўным значэнні спыняе яго.
Утварэнне падвойнага электрычнага слоя пры апусканні металу ў электраліт.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ПА́ЛІЎНЫ ЭЛЕМЕ́НТ,
хімічная крыніца току, у якой адбываецца непасрэднае пераўтварэнне хім. энергіі паліва ў электрычную; асн. частка электрахімічнага генератара.
Складаецца з 2 электродаў, раздзеленых цвёрдым (ці вадкім) электралітам. Паліва і акісляльнік падаюцца ў поласці, што мяжуюць з электродамі, і на паверхні раздзелу электрод — электраліт у прысутнасці каталізатараў (напр., плаціны) адбываюцца акісляльна-аднаўляльныя хім. рэакцыі (гл.Акісленне-аднаўленне). Электраліт забяспечвае прасторавае раздзяленне рэакцый акіслення і аднаўлення. У выніку рэакцый утвараюцца рознайменныя іоны, якія рэкамбінуюць у канчатковы прадукт рэакцыі, а на адпаведных электродах утвараецца лішкавы эл.зарад. Выкарыстоўваюцца як аўтаномныя крыніцы току ў сістэмах энергасілкавання касм. апаратаў, электрамабіляў, радыёапаратуры, радыёлакацыйных станцый і інш.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ПАЎПРАВАДНІКО́ВЫ ДЭТЭ́КТАРу ядзернай фізіцы,
прылада для рэгістрацыі іанізавальных выпрамяненняў, асн. элемент якой — крышталь паўправадніка.
Адчувальны слой П.д. — вобласць паўправадніка паблізу p-n-пераходу, якая абеднена носьбітамі току і мае высокае эл. супраціўленне. Іанізавальная часціца, што пранікла ў крышталь, генерыруе дадатковыя (нераўнаважныя) носьбіты току (электроны і дзіркі), у выніку чаго ў знешнім ланцугу ўзнікае імпульс току, які ўзмацняецца і рэгіструецца. Зарад, сабраны на электродах П.д., прапарцыянальны энергіі, выдзеленай часціцай пры праходжанні праз адчувальны слой. Можа працаваць як спектрометр, калі часціца поўнасцю тармозіцца ў ім. Выкарыстоўваюцца ў ядз. фізіцы, фізіцы элементарных часціц, а таксама ў хіміі, геалогіі, медыцыне і прам-сці.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЕ́ЙГЕР, Гайгер (Geiger) Ганс Іаган; 30.9.1882, г. Нойштат-ан-дэр-Вайнштрасэ, Германія — 24.9 1945),
нямецкі фізік-эксперыментатар. Чл. Берлінскай АН (1937). Вучыўся ў Эрлангенскім, Мюнхенскім і Манчэстэрскім ун-тах. З 1907 выкладаў у Манчэстэрскім, Кільскім, Цюбінгенскім, Берлінскім ун-тах. Навук. працы па атамнай і ядз. фізіцы, фізіцы касм. прамянёў. У 1908 вызначыў зарад электрона; разам з Э.Рэзерфардам вынайшаў прыладу для падліку асобных мікрачасціц (гл.Гейгераўскі лічыльнік); разам з англ. фізікам Дж.Неталам прапанаваў эмпірычную формулу сувязі пастаяннай распаду з энергіяй альфа-часціц (1911—12; закон Гейгера—Нетала); разам з В.Ботэ даказаў справядлівасць законаў захавання энергіі і імпульсу пры сутыкненнях элементарных часціц; разам з англ. фізікам Э.Марсдэнам эксперыментальна пацвердзіў формулу Рэзерфарда па рассейванні альфа-часціц.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КУЛО́НА ЗАКО́Н,
адзін з асн. законаў электрастатыкі, які вызначае сілу ўзаемадзеяння паміж двума кропкавымі зарадамі (гл.Зарад электрычны). Устаноўлены ў 1785 Ш.А.Кулонам і незалежна Г.Кавендышам (яго працы апублікаваны ў 1879) і з’яўляецца эксперым. абгрунтаваннем класічнай электрадынамікі.
Паводле К.з. 2 кропкавыя задачы q1 і q2 узаемадзейнічаюць у вакууме з сілай , модуль якой прама прапарцыянальны здабытку гэтых зарадаў і адваротна прапарцыянальны квадрату адлегласці г паміж імі: F = kq1q2/r2, дзе k = 1/4πε0, ε0 — электрычная пастаянная. Сіла накіравана ўздоўж прамой, што злучае зарады, і адпавядае прыцягненню рознаіменных зарадаў і адштурхоўванню аднайменных. Калі ўзаемадзейныя зарады знаходзяцца ў аднародным дыэлектрыку з дыэлектрычнай пранікальнасцю ε, сіла іх узаемадзеяння змяншаецца ў ε разоў. Абагульненне К.з. прыводзіць да Гаўса тэарэмы.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГА́ЎСА ТЭАРЭ́МА,
асноўная тэарэма электрастатыкі, якая ўстанаўлівае сувязь паміж патокам напружанасці эл. поля праз адвольную замкнёную паверхню і эл. зарадам, што знаходзіцца ўнутры гэтай паверхні. Вынікае з Кулона закону, устаноўлена К.Ф.Гаўсам (1839).
У інтэгральнай форме Гаўра тэарэма мае выгляд:
, дзе — паток вектара напружанасці эл. поля праз замкнёную паверхню S, q — зарад, абмежаваны паверхняй S, ε0 — электрычная пастаянная. Дыферэнцыяльная форма Гаўра тэарэмы:
, дзе — дывергенцыя вектара , ρ — шчыльнасць эл. зараду ў тым пункце прасторы, дзе вызначаецца . Гаўра тэарэма адно з Максвела ўраўненняў, адлюстроўвае той факт, што эл. зарады з’яўляюцца крыніцамі эл. поля; дазваляе вызначаць вектар пры зададзеным зарадзе (шчыльнасці зараду).
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЗАРА́ДУ ЗАХАВА́ННЯ ЗАКО́Н,
адзін з асн. законаў прыроды, паводле якога алгебраічная сума зарадаў электрычных замкнутай (эл. ізаляванай) сістэмы застаецца нязменнай пры любых працэсах унутры гэтай сістэмы. Устаноўлены ў 18 ст.
Адкрыццё электрона ў канцы 19 ст. і пратона ў пач. 20 ст. даказала, што эл.зарад звязаны з часціцамі (з’яўляецца іх унутранай уласцівасцю) і ёсць цэлы лік, кратны зараду элементарнаму. З.з.з. — вынік захавання колькасці часціц у тых працэсах, дзе няма іх узаемапераўтварэнняў. Напр., пры электрызацыі макраскапічных цел зараджаныя часціцы пераносяцца з аднаго цела на другое. У фізіцы элементарных часціц, для якой характэрна ўзаемапераўтварэнне часціц, нараджэнне «новай» часціцы суправаджаецца знікненнем «старой» з такім жа зарадам або нараджэннем яшчэ адной часціцы з зарадам процілеглага знака (напр., працэс нараджэння пары часціца—антычасціца).
адносіны магнітнага моманту элементарных часціц і сістэм, што складаюцца з іх (атамных ядраў, атамаў, малекул і інш.), да іх моманту імпульсу (мех. моманту).
М.а. для розных станаў атамных сістэм вызначаюцца формулай: γ = gγ0, дзе g — Ландэ множнік, γ0 — адзінка М.а. У выпадку арбітальнага руху электрона ў атаме γ0 = −e/2mec, для спіна электрона γ0 = −e/mec, для атамных ядраў γ0 = −e/2mpc, дзе e — элементарны эл.зарад, me і mp — маса электрона і пратона адпаведна, c — скорасць святла ў вакууме. М.а. вызначаюць ўздзеянне знешняга магн. поля на сістэму, якая мае магн. момант, і выкарыстоўваецца, напр., у квантавых магнітометрах з прэцэсіяй намагнічанасці мікрачасціц для вызначэння магн. індукцыі знешняга поля. Гл. таксама Лармара прэцэсія, Магнетон.