частка тэр.Памор’я, якая была ўключана ў склад Польшчы паводле Версальскага мірнага дагавора 1919. Вузкая (ад 30 да 200 км) паласа зямлі давала Польшчы выхад да Балт. мора, дзе ў сярэдзіне 1920-х г. быў пабудаваны порт Гдыня. Аддзяляў Усх. Прусію ад асн.тэр. Германіі, што было адной з прычын напружанасці ў германа-польскіх адносінах. У самой Польшчы да вызначэння «П.к.» ставіліся адмоўна. У сак. 1939 рэйхсканцлер Германіі А.Гітлер запатрабаваў ад польскага ўрада права будаўніцтва праз «П.К.» экстэрытарыяльных аўтастрад; адмова апошняга стала адной з зачэпак для германскай агрэсіі супраць Польшчы ў вер. 1939.
рускі музычны крытык, педагог. Скончыў Пецярбургскую кансерваторыю (1866), вучыўся ў А.Рубінштэйна. Праф. Маскоўскай (1867—86, з перапынкамі) і Пецярбургскай (1872—79) кансерваторый. З 1860-х г. выступаў у перыяд. друку. Аўтар цыкла артыкулаў пра творчасць М.Глінкі, П.Чайкоўскага, інш.рус. і зах.-еўрап. кампазітараў і артыстаў, па пытаннях муз. адукацыі, муз. навукі і інш. У шэрагу ацэнак выявіліся супярэчлівасць і кансерватызм яго эстэт. прынцыпаў: недаацэньваў творчасць кампазітараў «Магутнай кучкі», адмоўна ставіўся да новых тэндэнцый у музыцы 2-й пал. 19 ст. Выступаў і як літ. крытык.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ІАНІЗА́ЦЫЯ,
утварэнне дадатна або адмоўна зараджаных часціц (іонаў) і свабодных электронаў з эл. нейтральных атамаў ці малекул. Адбываецца ў рэчыве пад уплывам моцнага эл. поля, высокіх т-р, а таксама ад дзеяння ўдараў хуткіх зараджаных часціц і фатонаў у выніку таго, што атам ці малекула набывае лішкавую энергію, дастатковую для вылучэння электрона.
Пры І. газу з атамаў або малекул узнікаюць дадатны іон і свабодны электрон, якія застаюцца свабоднымі ці далучаюцца да інш. нейтральных атамаў або малекул, утвараючы комплексныя іоны. І. ў цвёрдых целах — пераход электронаў з атамаў асн. рэчыва ці з прымесных цэнтраў у зону праводнасці. Гл. таксама Зонная тэорыя, Электраправоднасць.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
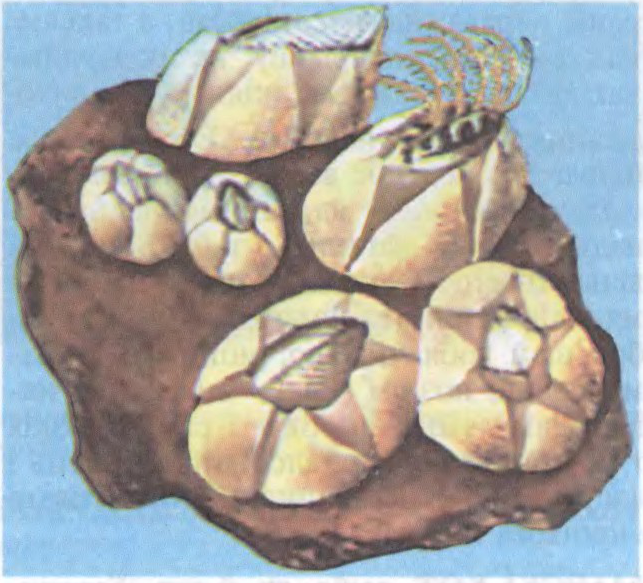
МАРСКІ́Я ЖАЛУДЫ́ (Balanomorpha і Venucomorpha),
2 надсямействы вусаногіх ракападобных жывёл падатр. тарацыкавых. Больш за 400 відаў. Пашыраны ва ўсіх акіянах да глыб. 160 м. Жывуць на дне, прымацоўваюцца да цвёрдага субстрату (скалы, канструкцыі) або да прадметаў ці жывёл (кіты, чарапахі і інш.), днішчаў суднаў (адмоўна ўплываюць на іх мараходныя якасці). Устойлівыя да неспрыяльных умоў асяроддзя, могуць некаторы час абыходзіцца без вады.
Вышыня вапністага пласціністага доміка-ракавіны, што ўкрывае цела, да 25 см, шыр. да 10 см. Кормяцца планктонам, водарасцямі, якія фільтруюць праз ракавіну з дапамогай 6 пар двухгалінастых ножак. Пераважна гермафрадыты, развіццё з лічынкай. Некат, ядомыя. Гл. таксама Вусаногія.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БІЯТО́П (ад бія... + грэч. topos месца),
участак зямной паверхні (сушы або вадаёма) з аднатыпнымі абіятычнымі ўмовамі асяроддзя (рэльеф, глебы, клімат і інш.), які займае вызначаны біяцэноз; неарган. кампанент біягеацэнозу. Характэрны для дадзенага біятопа комплекс умоў вызначае відавы склад арганізмаў, асаблівасці іх існавання, трапляе пад уплыў біяцэнозу і мяняецца ад яго ўздзеяння.
Для фізіка-геагр. умоў Беларусі характэрны біятопы: балоты (нізінныя, пераходныя, вярховыя), лясы (хваёвыя, яловыя, альховыя, бярозавыя і інш.), лугі (заліўныя, сухадольныя), палі, прэсныя воды (рэкі, азёры, вадасховішчы). Пераўтваральная дзейнасць чалавека на біятоп можа прывесці да непажаданых змен у біягеацэнозах ці поўнага іх разбурэння, што адмоўна ўплывае на склад і размеркаванне фауны і флоры. Падобныя біятопы аб’ядноўваюць у біяхоры, сукупнасці якіх складаюць біяцыклы.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЕ́ЛЬФАНД (Хаім Янкель Шыманавіч) (літ.псеўд.А.І.Літвак; 1877, г. Слуцк Мінскай вобл. — 1932),
палітычны дзеяч і публіцыст. Адзін з кіраўнікоў Бунда, з 1905 чл. яго ЦК. Неаднаразова быў арыштаваны, у 1896—1902 у ссылцы ў Якуцку. У 1915—17 прадстаўнік Бунда ў ЗША. Пасля Лют. рэвалюцыі 1917 вярнуўся ў Расію. Да Кастр. рэвалюцыі 1917 паставіўся адмоўна. На XI канферэнцыі Бунда (1919, Мінск) узначаліў с.-д. крыло. У 1921 арыштаваны органамі ДПУ, пасля вызвалення жыў у Вільні. З 1925 у ЗША.
Тв.:
Мелкобуржуазный социализм на еврейской почве: (Критика «теории» социалистов-сионистов). СПб., 1906;
Поалей сионизм: Новое течение в рус. еврействе — рабочий сионизм: Критический очерк. СПб., 1907.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КІСЛО́ТНЫЯ ДАЖДЖЫ́, кіслыя дажджы,
атмасферныя ападкі, забруджаныя вокісламі азоту і серы, што трапляюць у паветра з выкідамі прамысл. прадпрыемстваў. Выпадаюць у выглядзе раствораў кіслот, якія ўтвараюцца пры ўзаемадзеянні азотнага і сернага ангідрыдаў з атм. вадой. Адмоўна ўплываюць на стан навакольнага асяроддзя: павялічваюць кіслотнасць глебы, аказваюць біяцыднае ўздзеянне на жывыя арганізмы, пагражаюць здароўю чалавека, шкодзяць прамысл. і арх. пабудовам. Могуць выпадаць на значнай адлегласці ад крыніц забруджвання. Колькасць К.д. і іх адмоўны ўплыў на навакольнае асяроддзе штогод павялічваюцца. Памяншэнне ці спыненне выкідаў у паветра прамысл. прадпрыемствамі шкодных рэчываў, у т. л. вокіслаў азоту і серы, правядзенне інш. мерапрыемстваў па ахове атмасферы дапамагаюць зменшыць кіслотнасць атм. ападкаў, спрыяюць спыненню К.д.
аўстрыйскі музыказнавец. Вучыўся ў Пражскім і Венскім (з 1856 выкладаў у ім) ун-тах. Трактатам «Аб музычна-прыгожым» (1854) пачаў дыскусію аб праграмнай і «чыстай» («абсалютнай») музыцы. Яго асн. ідэя — «муз. змест ёсць рух гукавых формаў» — сцвярджала муз. мастацтва як асобую, ізаляваную галіну духоўнай дзейнасці, пры тлумачэнні якой не прыдатныя немуз. асацыяцыі. Крытыкаваў палажэнне афектаў тэорыі і погляды франц. асветнікаў, што лічылі вызначальным пачаткам муз. выразнасці інтанацыі чалавечай мовы. Прынцыповы прыхільнік інструментальнай музыкі (у асн. І.С.Баха, венскіх класікаў, І.Брамса), адмоўна выказваўся аб рамантычнай праграмнай і опернай музыцы, пераважна Ф.Ліста і Р.Вагнера. Яго працы і крытычная дзейнасць стымулявалі паглыбленае вывучэнне ўнутр. заканамернасцей музыкі, у т. л. стварэнне тэорыі муз. стыляў Т.Рымана і Т.Адлера.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БУ́ФЕРНАСЦЬ ГЛЕ́БЫ,
уласцівасць глебы перашкаджаць змяненню сваёй актыўнай кіслотнасці (pH) пры ўздзеянні кіслот або шчолачаў.
Абумоўлена наяўнасцю ў ёй калоідаў, якія маюць здольныя да абмену іоны: іоны вадароду вызначаюць буфернасць у адносінах да шчолачаў, а іоны асновы — да кіслот. Глебавы раствор валодае буфернай уласцівасцю, калі ў ім ёсць солі моцнай асновы (натрыю, калію, кальцыю) і слабых, пераважна арган. кіслот (гумінавай, вугальнай і інш.), у сумесі са слабай кіслатой, у якой ёсць агульны з соллю аніён. Буфернасць глебы залежыць звычайна ад калоіднай і глеістай фракцый глебы. Найб. буферныя багатыя гумусам глебы цяжкага грануламетрычнага складу: чарназёмныя, тарфяністыя і інш. Расліны адмоўна рэагуюць на рэзкія ваганні pH глебы, таму буфернасць глебы адыгрывае вял. ролю ў іх росце і развіцці; яе можна павысіць унясеннем арган. ўгнаенняў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КАНТА́КТНАЯ РО́ЗНАСЦЬ ПАТЭНЦЫЯ́ЛАЎ,
рознасць патэнцыялаў паміж праваднікамі з рознай работай выхаду, якія знаходзяцца ў электрычным кантакце, ва ўмовах раўнавагі тэрмадынамічнай.
Пры кантакце двух праваднікоў паміж імі адбываецца абмен электронамі праводнасці. У выніку праваднік з меншай работай выхаду зараджаецца дадатна, з большай — адмоўна. У вобласці кантакту ўзнікае эл. поле, накіраванае так, што патокі электронаў у абодвух напрамках ураўнаважваюцца, і ўстанаўліваецца пастаянная К.р.п. Яна роўная рознасці работ выхаду праваднікоў. аднесенай да зараду электрона. К.р.п. залежыць ад прыроды правадніка, стану яго паверхні і можа дасягаць некалькіх вольт. На існаванні К.р.п. грунтуецца работа важнейшых элементаў паўправадніковай электронікі: p—n пераходаў і кантактаў метал—паўправаднік; яна выкарыстоўваецца для прамога пераўтварэння цеплавой энергіі ў электрычную; яе трэба ўлічваць пры канструяванні электравакуумных прылад.