Беларуская Савецкая Энцыклапедыя (1969—76, паказальнікі; правапіс да 2008 г., часткова)
Verbum
анлайнавы слоўнікБеларуская Савецкая Энцыклапедыя (1969—76, паказальнікі; правапіс да 2008 г., часткова)
Беларуская Савецкая Энцыклапедыя (1969—76, паказальнікі; правапіс да 2008 г., часткова)
ГАРА́ЧАЎ (Іван Іванавіч) (
Тв.:
Пути решения проблемы
Кормление высокопродуктивных коров.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ІВАНО́Ў (Вадзім Ціханавіч) (
расійскі хімік-арганік.
Тв.:
Путь к синтезу

Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГАЛУ́ШКА (Васіль Міхайлавіч) (
Тв.:
Рациональное использование протеина // Пути решения проблемы
Питательность кормов: Справ. пособие.
Приготовление кормов для свиней.

Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БІЯСІ́НТЭЗ (ад бія... + сінтэз),
утварэнне ў жывых арганізмах складаных
Пачатковыя пастаўшчыкі энергіі для біясінтэзу — зялёныя расліны і фотасінтэзавальныя бактэрыі, што акумулююць сонечную энергію (
Біясінтэз можа быць ажыццёўлены і ў
Літ.:
Биосинтез
Молекулярная биология клетки:
Ленинджер А. Основы биохимии:
А.М.Ведзянееў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БЯЛКІ́,
прыродныя высокамалекулярныя
Агульны
Літ.:
Бохински Р. Современные воззрения в биохимии:
Ленинджер А. Основы биохимии:
Гершкович А.А. От структуры к синтезу
Овчинников Ю.А. Химия жизни: Избр.
У.М.Рашэтнікаў.
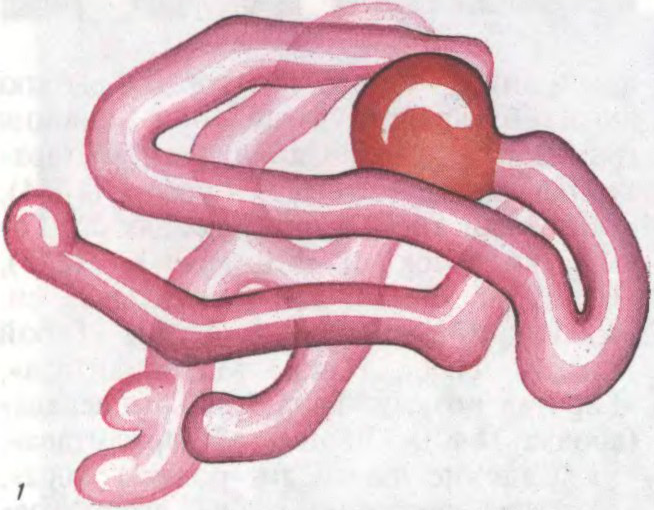
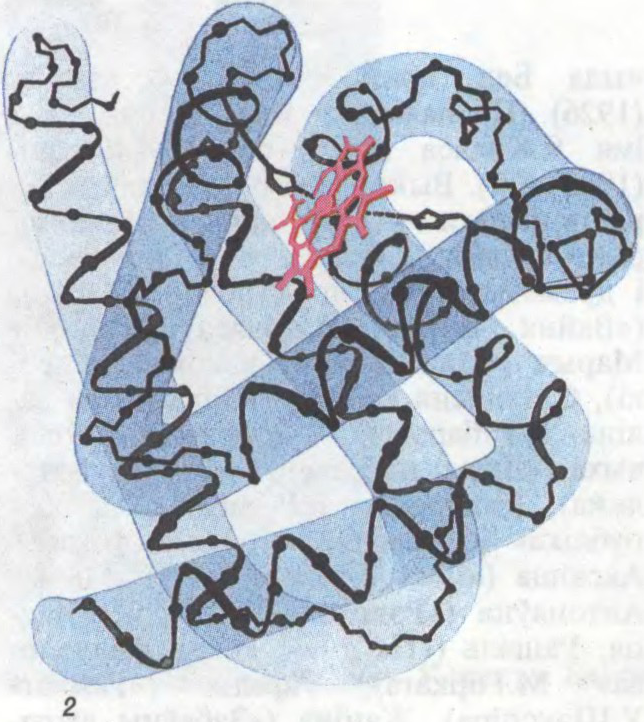
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЗАСЛУ́ЖАНЫ БУДАЎНІ́К РЭСПУ́БЛІКІ БЕЛАРУ́СЬ,
ганаровае званне, якое прысвойваецца высокапрафес. работнікам будаўніцтва і будаўнічай індустрыі, якія працуюць у будаўнічых,
Заслужаныя будаўнікі Рэспублікі Беларусь
1960. У.Ц.Атрошчанка, Е.С.Галаўнёва, С.А.Грыгор’еў, В.Ф.Дзем’янюк, А.А.Дзігілевіч, І.А.Доўнар, А.А.Жалняровіч, Р.С.Забалееў-Ізотаў, Я.Ф.Казлоўская, З.А.Караткевіч, І.К.Колычаў, А.С.Лапуноў, М.Х.Ломскі, У.П.Маскалёў, В.І.Свецікаў, В.А.Слепухоў, П.М.Сцяблюк, Ц.К.Фандо, Я.Ф.Філічонак, П.Г.Фядоценка, І.Э.Шаблоўскі, В.Ш.Шайхлісманаў, С.І.Ярашэвіч.
1961. А.В.Астроўскі, У.М.Вараксін, В.К.Гаравой, А.М.Громаў, М.К.Гуд, І.К.Дуброўскі, Г.У.Заборскі, А.Ц.Маграшоў, В.Дз.Макавец, Т.М.Макараў, Л.А.Новікава, І.А.Паддубіцкі, М.І.Паўлаў, М.М.Сівухін, М.П.Тарасенка, І.Дз.Шлапакоў.
1962. Р.А.Абразцова, А.С.Абухоўскі, Я.І.Анісімаў, Ф.Ф.Арцёменка, С.С.Атаеў, П.Л.Бабак, А.Г.Базаеў, С.С.Батурын, М.Н.Богуш, К.А.Бондараў, Л.С.Булах, М.Л.Вараб’ёў, І.П.Ведзмедзь, А.П.Воінаў, М.С.Галуза, Г.М.Глезер, І.І.Грыгаровіч, Л.А.Жук, І.М.Жыжаль, У.В.Казюля, А.Н.Каралёў, У.А.Кароль, М.В.Кашчэеў, Н.І.Лісавец, К.А.Магілеўскі, А.С.Мазейка, В.А.Мароз, У.Г.Міхайлаў, А.Ф.Мураўскі, Л.С.Паперна, Г.П.Паўлоўскі, В.Л.Пятніцкі, І.М.Рабцаў, Р.І.Рубанаў, М.А.Свірскі, М.П.Семянцоў, Г.І.Страіцелеў, В.А.Талмачоў, С.М.Траногін, В.Л.Трэйда, В.М.Фамін, М.А.Хомчанка, В.Я.Цвяркун, П.П.Цімашэнка, К.П.Цюцюноў, М.В.Шчэпкін, І.А.Юркоў.
1963. А.В.Анцімонаў, М.Ц.Архіпец, Л.М.Ашмян, В.С.Бабкоў, Ф.Ц.Бажок, А.У.Байцова, І.С.Бахір, Я.Е.Гайко, Я.П.Гоцін, А.П.Гушча, К.У.Джыгера, Г.Я.Жолудзеў, В.П.Зенін, А.С.Крэмень, А.В.Круль, М.С.Кускоў, В.С.Левін, М.Ф.Майдановіч, Л.П.Мацкевіч, М.М.Машкін, У.М.Мудры, С.І.Мядзведскі, В.Ф.Наймушын, П.М.Новікаў, Я.Ф.Паўленка, А.Ш.Плоткін, Я.А.Раговін, М.З.Санкін, Г.І.Татурын, А.Б.Уласевіч, К.Дз.Цвярдоў, І.М.Чарненка.
1964. І.А.Віленскі, М.Дз.Галубенкаў, М.В.Кавалёў, І.І.Корсакаў, В.С.Крывіцкі, Р.І.Мілановіч, А.В.Пруднікаў, У.М.Рубанік, В.Ф.Хлімянцоў, Л.С.Храмцоў.
1965. А.Б.Бандарэнка, П.І.Бязручкін, А.К.Ворса, І.М.Вятрук, І.Ц.Гапеенка, М.Я.Глушанкоў, М.К.Дзямешка, Ф.І.Дудараў, У.М.Жур, А.І.Кавалеўскі, М.У.Конік, Ф.Я.Крот, Ц.М.Кушнераў, В.С.Ларын, Р.І.Лебедзеў, У.Я.Ліцкевіч, П.С.Лукашук, А.Дз.Майсяевіч, М.Я.Мананкоў, Б.М.Марагоўскі, В.Г.Маршалаў, М.С.Медзвядзенка, Г.К.Мітраховіч, Дз.Ф.Міхайленка, П.М.Наваржэўскі, П.Ф.Пацей, А.А.Пількевіч, С.М.Раўкоў, Я.Ф.Савіцкі, І.С.Салей, С.І.Серафімовіч, І.Х.Сіманоўскі, М.В.Скрабкоў, Л.С.Собаль, А.П.Старавойтаў, І.І.Сямак, К.М.Тандзітнікаў, М.Г.Ушкевіч, П.І.Чарнавусаў, В.П.Чарнышоў, М.І.Шалдыга, Б.А.Шаршнёў, Г.П.Шмігельскі, М.І.Шымель, І.П.Янкоўскі, М.П.Ярашэвіч.
1966. В.Е.Алексяюк, К.А.Бародзіч, І.М.Бірыла, К.І.Вагін, Л.Я.Васілеўскі, Г.А.Вялічка, А.М.Гершаў, Г.С.Дашын, В.Р.Жылін, А.С.Зуб, С.Ф.Кажамякін, М.П.Кажан, П.І.Клімук, І.К.Лазарэнка, А.К.Лісоўскі, К.С.Мілашэўскі, А.Ф.Мітраховіч, Д.П.Надгачаеў, Р.Я.Паламарчук, Л.В.Парыйскі, Я.П.Ракомса, С.А.Сапун, П.Ф.Філіпаў, У.А.Хадорык, Ф.І.Цімафееў, С.Я.Цярохін, Л.К.Цяцерын, А.С.Чмыр, І.М.Шараеў, І.К.Юркоў.
1967. І.С.Вараб’ёў, М.П.Варэнцаў, Я.І.Дрозд, А.А.Кавалевіч, М.М.Кавалевіч, Е.А.Каўгарэна, Ц.І.Кузьмін, А.Дз.Лакіда, М.В.Леаненка, Ф.Ф.Леановіч, С.І.Шульга.
1968. П.Р.Аўсянікаў, М.І.Бадзько, Г.С.Бандарэнка, К.В.Варон ша, А.К.Васілеўскі, Л.Г.Гаўрылаў, Р.Я.Герцовіч, В.П.Глявіцкі, А.А.Грабеннікаў, М.А.Грыгарук, П.Р.Дадонаў, М.Г.Дзівакоў, А.Я.Каржуеў, М.К.Кесель, Ф.В.Магіленка, К.Я.Масько, М.І.Мацэль, М.М.Мядзведскі, І.Б.Палінскі, М.І.Пісарчык, С.Ф.Пятроў, В.Дз.Сабалеўскі, І.М.Сак, А.З.Сацікаў, А.І.Хамянкоў, К.С.Цывінскі, Ф.М.Шакай, Ф.І.Шчасны, Ф.І.Юшкевіч, В.Н.Ярковіч.
1969. Дз.Д.Атрашкевіч, Ю.К.Гарашчук, Г.М.Губчык, М.М.Епіфанаў, М.І.Касценіч, С.Ц.Кірыльчук, М.І.Кудзелін, А.В.Кулікоўскі, Б.С.Курлена, І.В.Кушнераў, Ф.С.Маскоў, В.М.Няхаеў, П.Р.Паўлоўскі, А.М.Печкуроў, Дз.Дз.Прашчаеў, У.М.Рыжыкаў, С.П.Самуха, М.П.Сінцова, М.М.Унукоўскі, І.І.Ціханаў, А.А.Чыжык, С.М.Шахно, К.С.Якімовіч.
1970. П.Н.Антоненка, М.І.Бандзюк, С.С.Бычкоўскі, І.І.Валадашчык, А.Ф.Гаўрысёў, М.І.Герасімчук, Д.Ц.Гузман, Ю.Р.Іваноў, С.М.Ізотаў, І.П.Кавалёў, С.В.Казакоў, Л.В.Казлоў, В.І.Кандраценка, М.Л.Каралюк, Р.В.Куляеў, Л.І.Кунічкіна, М.Р.Лукашэнка, Р.І.Ляшэнка, К.П.Малькевіч, С.І.Маршчэнка, М.М.Маталыцкі, М.Ф.Мацюк, А.І.Наканечны, Р.Л.Пабядзінскі, У.В.Пакацілаў, В.І.Пацюк, П.Я.Пісарчык, М.А.Пісарык, Ф.П.Пыжык, А.А.Раманоўскі, А.А.Рамашэўскі, Д.Я.Родаў, М.Ц.Свінко, Р.С.Сілівончык, М.В.Сярмяжка, А.А.Ларасаў, А.А.Фурсевіч, Л.У.Чарапко, М.А.Шлойда, М.С.Шумакоў.
1971. І.А.Багданаў, Р.І.Бандарэнка, М.А.Баранаў, Л.В.Голубева, А.К.Іваноў, Ф.І.Кажарскі, І.І.Казлоўскі, С.І.Казлюк, Б.М.Кіціс, Р.Дз.Купрыенка, Ф.А.Лапуноў, М.А.Літоўчанка, В.І.Луцык, М.Н.Маліноўскі, А.Н.Мінаеў, У.У.Мінкевіч, К.К.Петрушкевіч, М.І.Пунько, Г.М.Рагачова, І.В.Сабалеўскі, П.М.Сідараў, А.І.Сініцын, М.К.Смоліч, А.Л.Трубяцкой, А.А.Трухан, М.А.Федарчук, А.І.Цыганок, М.І.Яскевіч.
1972. Ф.А.Андросік, М.Д.Бараболкін, П.Л.Бароўскі, В.І.Барысенка, Б.А.Бразінскі, Я.І.Грыняк, М.М.Грышкавец, А.С.Карамышаў, В.С.Карэўка, А.К.Клявін, В.І.Кувашоў, У.І.Курашоў, П.І.Лаўрэнка, У.А.Малашэвіч, У.Ф.Маціеўскі, Л.У.Парахонька, І.М.Пацярайчанка, У.Д.Рабянок, В.А.Руды, А.Я.Фамін, В.М.Чаркін, С.В.Шапавалаў, Р.Ф.Шукаў, Я.С.Юшкоўская, С.А.Язэпчык.
1973. А.С.Арцюх, В.В.Дзеўнер, А.П.Забела, А.С.Закоршмены, П.П.Красько, М.В.Крутаў, Г.Н.Кудрашоў, С.С.Макавецкі, Г.П.Навасельскі, В.П.Русак, М.М.Рыгачэвіч, М.А.Сілін, П.М.Сябекін, В.А.Хворык, А.М.Чыгрэцкі.
1974. Я.В.Барысевіч, М.У.Бегуноў, М.М.
1975. А.П.Арцем’еў, Б.С.Бальцэвіч, М.Ф.Барадулька, І.Ц.Бялоў, К.А.Валенцік, К.А.Вяранчык, А.І.Вярстак, У.Е.Гарбацкі, Г.С.Друкараў, І.М.Еўдамашчанка, Г.П.Карлаў, І.Я.Качанаў, А.М.Крапіўка, Р.Р.Кучук, М.А.Мяжэвіч, М.П.Муранаў, П.І.Панамарэнка, У.В.Пішчыкаў, І.С.Рабочы, І.І.Раманееў, І.К.Рубахаў, В.Ц.Саяпін, І.П.Севасцьянаў, Б.А.Халадцоў, А.Ф.Шалудзько, А.С.Шчарбінін, В.А.Шыманскі, А.М.Яўціхіеў.
1976. А.І.Акатаў, А.М.Аксёнаў, У.Ц.Алізарэвіч, П.С.Амоненка, У.А.Андросік, К.Дз.Багданаў, С.Ц.Баярчук, Н.С.Бельчанка, П.П.Бондараў, А.А.Боршч, П.А.Брэль, Р.С.Бут-Гусаім, М.А.Гарбузаў, В.Ф.Вішнеўскі, І.А.Гапонаў, У.Ц.Гніза, Л.М.Голубева, У.С.Дзятлаў, А.Ц.Драмянкоў, В.В.Дрэнь, С.М.Жалубоўскі, А.С.Зданевіч, М.Л.Зеліхоўскі, У.І.Касоўцаў, А.Т.Касцюкоў, В.Ф.Касько, М.І.Кісялёў, Я.С.Костусеў, А.І.Краўчанка, І.А.Мейлах, Б.М.Мілер, І.М.Наваселецкі, Ф.А.Назарчук, М.М.Несцераў, І.Р.Пісарук, У.А.Рагачоў, А.У.Сарокін, І.Д.Страліцкі, А.М.Туз, Я.А.Шушкевіч, Я.К.Шэрыкаў.
1977. М.М.Грышчэня, Р.М.Дарашэнка, А.Н.Каўрэй, А.А.Корзун, М.Ф.Лунін, М.У.Насулька, П.Л.Пастушэнка, Л.У.Лабалевіч, М.К.Тышкевіч, Б.Р.Ханін, М.Р.Чумакоў, У.І.Шылаў.
1978. В.І.Альхоўскі, П.В.Баравікоў, А.В.Богуш, В.М.Куракевіч, А.А.Луцык, Дз.Ф.Махавікоў, Р.Ф.Сялевіч, У.К.Шульга, К.С.Шулянчык, П.М.Шыпаў.
1979. І.А.Антановіч. Э.В.Аўчыннікаў, А.І.Барташ, І.М.Бутрымовіч, А.А.Дунаеў, М.Н.Зельдас, Л.Б.Любезны, І.І.Ляўко, М.М.Мартынаў, Я.І.Маўчан, І.А.Салодкі, І.П.Чорствы.
1980. М.А.Арэф’еў, М.К.Балыкін, У.І.Бобр, М.М.Верамеюк, М.С.Гапееў, Дз.І.Гвоздзеў, Я.М.Гопша, А.Ц.Доўгі, І.І.Дубовік, І.А.Кенік, А.Т.Кічкайла, А.Т.Кобля, В.А.Крамар, М.М.Лавянецкі, І.Р.Ладуцька, К.А.Лапец, М.М.Лапко, Ф.Р.Літвіновіч, Г.А.Мадалінскі, В.І.Макараў, Я.А.Мартыненкаў, Р.М.Махліс, В.В.Пыжык, К.П.Пятрыніч, ф.Ф.Раманчук, Ю.А.Сабалеўскі, М.Ц.Салдаткін, А.К.Самаросенка, М.П.Сачок, М.П.Свірыдовіч, П.В.Селівонік, Я.Г.Селівончык, А.А.Сідарук, А.Г.Танаян, Ф.Я.Таўкачоў, В.І.Трубянок, І.У.Філоненка, У.І.Харужы, А.М.Целеш, З.М.Шалыга, І.Р.Шкрабнёў.
1981. М.П.Аляксюк, У.І.Газдзецкі, Л.Ц.Кацько, А.І.Малыха, М.М.Малярэвіч, М.Я.Талсташэеў, М.С.Трусевіч, Г.В.Юшкевіч, М.Я.Юшкевіч, М.Я.Ярошкін.
1982. Р.М.Баглай, У.М.Гаўрылаў, І.І.Дзянішчык, В.І.Жалудко, Л.І.Максімовіч, З.Я.Паланецкі, М.Ф.Сайкоўскі, Г.Т.Сіўцоў, І.А.Тарасавец, А.В.Тарасевіч, А.Р.Цыбуля, В.М.Шантаровіч, П.У.Якубчык.
1983. Я.П.Стойка.
1984. М.Д.Белы, А.З.Бялевіч, В.М.Веліканаў, А.П.Дзямчук, Д.Д.Дук, А.С.Емяльянаў, М.М.Кандакоў, М.Я.Краснарадцаў, І.І.Лазараў, Ю.А.Пуплікаў, А.І.Смольскі, М.І.Тарун.
1985. Я.Р.Брычкоўскі, В.І.Бялоў, А.Г.Ваўранюк, В.М.Ветраў, Я.Ф.Вінакураў, Дз.П.Дземчанка, В.С.Зданевіч, І.П.Капітула, А.А.Каспяровіч, М.І.Папоў, П.П.Пракаповіч, Ф.М.Русак, М.С.Такоеў, Я.А.Траян, А.Дз.Фядотаў, А.В.Цадзілка, П.М.Ціханкоў.
1986. У.В.Аляшкевіч, Я.С.Бельская, Г.К.Варакса, В.І.Кажураў.
1987. І.З.Башмакоў, В.С.Барысевіч, А.У.Князеў, А.П.Кучэйка, У.М.Лагута, К.Л.Пятроўскі, А.Л.Сазонаў, П.К.Чыгірык.
1988. У.І.Казлоўскі, І.А.Нялюбін, М.С.Шамес.
1989. М.М.Дабрыцкі, М.Е.Капытоў, Ю.Б.Колакалаў, Л.І.Рыжкоў, У.С.Тамашук, А.Т.Шота, І.П.Шпорта, Ю.А.Шэпелеў.
1990. Ю.С.Астравух, Г.А.Баранаў, В.П.Баліцкі, Ф.Я.Бойка, М.К.Звонкін, Б.І.Касцюкевіч, В.А.Кулакоўскі, А.А.Ламакаў, В.І.Міцкевіч, І.І.Свярдлоў, В.А.Смірноў, Г.І.Станчык, В.В.Цімохін, П.М.Ціхановіч, А.Ф.Чуясаў, Л.С.Штырыкаў.
1991. М.З.Ашэйчык, В.Ф.Залога, В.В.Каржэўскі, Л.І.Мароз, Л.Ф.Марозаў, А.Л.Міхалевіч, У.П.Набокаў, В.П.Пятрыніч, А.Ф.Усціновіч, М.Дз.Цюшко.
1993. П.Дз.Андрэйкавец, А.П.Гапееў, Л.М.Красуцкі, С.А.Мазур, А.Я.Піскоўскі, К.М.Пячко, Л.Л.Чайлытка, А.Я.Янчэўскі.
1994. А.Дз.Зарубін, Р.Б.Кацынель, С.К.Лемеш, Т.І.Паляшчук, Л.І.Тарашкевіч, Ю.І.Уладзіміраў, У.І.Францкевіч, А.В.Якубёнак.
1995. Я.М.Галубкоў, Л.Б.Ваўчэцкі, А.У.Луцко, А.М.Макараў, У.Ц.Рагель, В.П.Сянцюраў.
1996. І.Я.Галіноўскі, С.У.Кунцэвіч, С.У.Міхалёнак.
1997. І.Р.Гапееў, В.А.Кулеба, Р.А.Шпакоўскі, А.М.Якаўлеў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)