КІНАРЫ́НХІ (Kinorhyncha),
клас марскіх першаснаполасцевых чарвей. 2
Даўж. 0,1—1
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КІНАРЫ́НХІ (Kinorhyncha),
клас марскіх першаснаполасцевых чарвей. 2
Даўж. 0,1—1
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КІНІ́НЫ (ад
фізіялагічна актыўныя поліпептыды: брадыкінін, лізілбрадыкінін і метыяніллізілбрадыкінін. У жывёл утвараюцца ў плазме крыві ці міжклетачных прасторах пры расшчапленні неактыўнага папярэдніка — кінінагена. Рэгулююць
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КЛАДО́ФАРА (Cladophora),
род зялёных водарасцей
Макраскапічныя водарасці ў выглядзе рыхлых дзярнінак, шчыльных паўшарападобных калоній ці бясформенных скопішчаў цёмна-зялёных нітак. Прымацаваны да субстрату рызоідамі або плаваюць свабодна, утвараючы ціну. Ніткі галінастыя, з адным радам выцягнутых шмат’ядравых
Т.М.Міхеева.

Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДЭТЭРМІНА́ЦЫЯ (ад
А.С.Леанцюк.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЗІМАЎСТО́ЙЛІВАСЦЬ
здольнасць раслін процістаяць маразам, рэзкім ваганням т-ры, высушванню, выправанню і
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ІНТЭРФЕРО́Н,
бялок, што ўтвараецца ў клетках арганізмаў млекакормячых і птушак пры вірусных інфекцыях; неспецыфічны фактар супрацьвіруснага імунітэту. Малекулярная маса 25000—110000. Асаблівасць І. — высокая відавая спецыфічнасць (
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
МАКРАЭЛЕМЕ́НТЫ,
хімічныя элементы, што ўтрымліваюцца ў тканках жывёльных і раслінных арганізмаў у адносна высокіх канцэнтрацыях (10−2% і вышэй). Да М. адносяцца кісларод, вадарод, вуглярод, азот, кальцый, фосфар, натрый, калій, сера, хлор, магній; з іх утвораны неарган. (
М.К.Кеўра.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
МЕНДЭЛІ́ЗМ,
вучэнне аб заканамернасцях атрымання ў спадчыну адзнак арганізма. Распрацаваны Г.Мендэлем (1865). Адыграў
Літ.:
Гайсинович А.Е. Восприятие менделизма в России и его роль в развитии дарвинизма // Природа. 1982. № 9.
Р.Г.Заяц.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
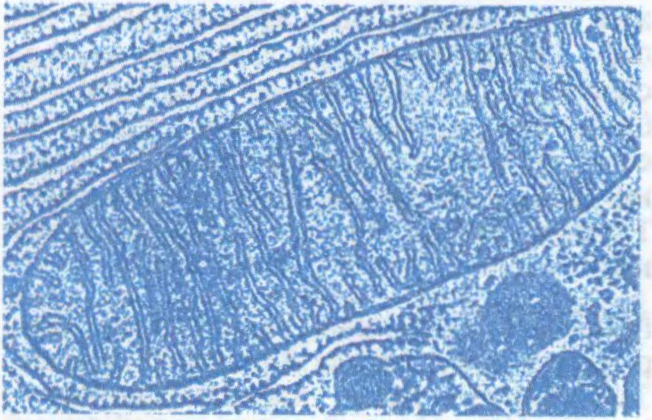
МІТАХО́НДРЫЯ (ад
арганоід
А.С.Леанцюк.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГІСТАЛО́ГІЯ (ад гіста... + ...логія),
навука, якая вывучае заканамернасці развіцця, будову і жыццядзейнасць тканак і органаў жывёл і чалавека. Даследуе комплексы
Развіццё гісталогіі як
На Беларусі развіццё гісталогіі звязана з арганізацыяй кафедры гісталогіі ў
Літ.:
Гистология. 4 изд.
Голуб Д.М. Очерки развития морфологии в
А.С.Леанцюк.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)