надатрад прамянёвапёрых рыб. Каля 40
Даўж. ад 7
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
надатрад прамянёвапёрых рыб. Каля 40
Даўж. ад 7
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БІЯЛАГІ́ЧНЫ ПРАГРЭ́С,
эвалюцыйны ўздым дадзенай групы арганізмаў у барацьбе за існаванне, які характарызуецца
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КІСЦЯПЁРЫЯ (Crossopterygii),
выкапнёвыя
Даўж. выкапнёвых форм ад 7
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АСЕМЯНЕ́ННЕ,
працэс, які забяспечвае ў жывёл сустрэчу палавых клетак — яец і сперматазоідаў; папярэднічае апладненню. Поспех асемянення забяспечваецца адначасовым выспяваннем гамет абодвух палоў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ОСМАРЭГУЛЯ́ЦЫЯ (ад осмас +
сукупнасць
А.С.Леанцюк.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
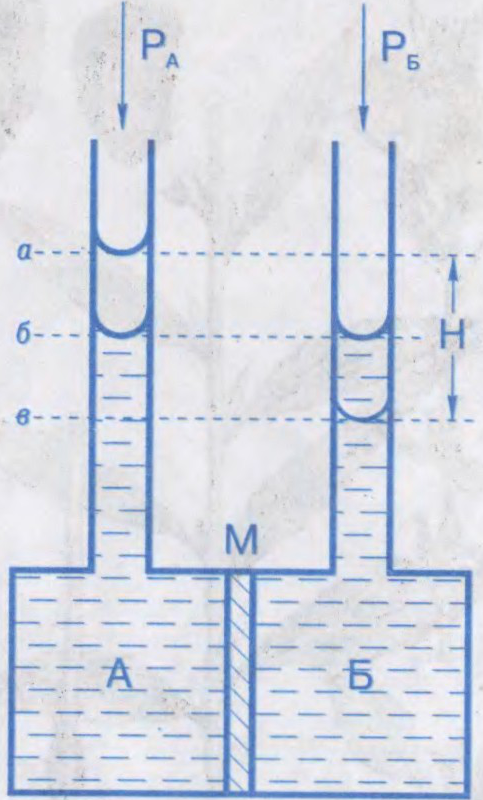
АСМАТЫ́ЧНЫ ЦІСК,
лішкавы гідрастатычны ціск раствору, які перашкаджае дыфузіі растваральніку праз паўпранікальную перагародку; тэрмадынамічны параметр. Характарызуе імкненне раствору да зніжэння канцэнтрацыі пры сутыкненні з чыстым растваральнікам пры сустрэчнай дыфузіі малекул растворанага рэчыва і растваральніку. Абумоўлены змяншэннем хімічнага патэнцыялу растваральніку ў прысутнасці растворанага рэчыва. Роўны лішкаваму вонкаваму ціску, які неабходна прыкласці з боку раствору, каб спыніць осмас. Вымяраецца ў паскалях.
Вымярэнні асматычнага ціску пачаў у 1877
Асматычны ціск адыгрывае важную ролю ў жыццядзейнасці жывых клетак і арганізмаў. У клетках і
Літ.:
Курс физической химии.
Пасынский А.Г. Коллоидная химия. 3 изд.
Гриффин Д., Новик
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
МЕЗАЗО́ЙСКАЯ ЭРАТЭ́МА (Э́РА),
мезазой
[ад мез(а)... +
другая эратэма слаёў фанеразою, якая ў агульнай стратыграфічнай шкале знаходзіцца паміж палеазойскай эратэмай (эрай) і кайназойскай эратэмай (эрай), і адпаведная ёй эра
У трыясавым перыядзе ў многіх абласцях Зямлі адбылося павелічэнне
Марскія геасінклінальныя адклады прадстаўлены пясчанікава-сланцавымі, часам з эфузівамі, і вапняковымі тоўшчамі; платформенныя — пясчана-гліністымі пародамі з глаўканітам і праслоямі фасфарытаў, на
На Беларусі вядомы адклады ўсіх 3 сістэм М.э. (э.).
В.С.Акімец.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)