Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДУБІ́НІН (Мікалай Пятровіч) (н. 4.1.1907, г. Кранштат Ленінградскай вобл., Расія),
савецкі генетык. Акад.АНСССР (1966; чл.-кар. 1946). Чл. шэрагу замежных АН. Скончыў Маскоўскі ун-т (1928). З 1932 працаваў у шэрагу н.-д. устаноў АНСССР, у 1966—81 дырэктар Ін-та агульнай генетыкі АНСССР. Адкрыў (разам з А.С.Сераброўскім) падзельнасць гена. Даследаваў эфект становішча гена, пытанні генетыкі папуляцый. Навук. працы па эвалюцыйнай, радыяцыйнай і касм. генетыцы, тэарэт. асновах селекцыі. Ленінская прэмія 1966.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АЛЕ́ЛІ (ад грэч. allēlōn адзін аднаго, узаемна),
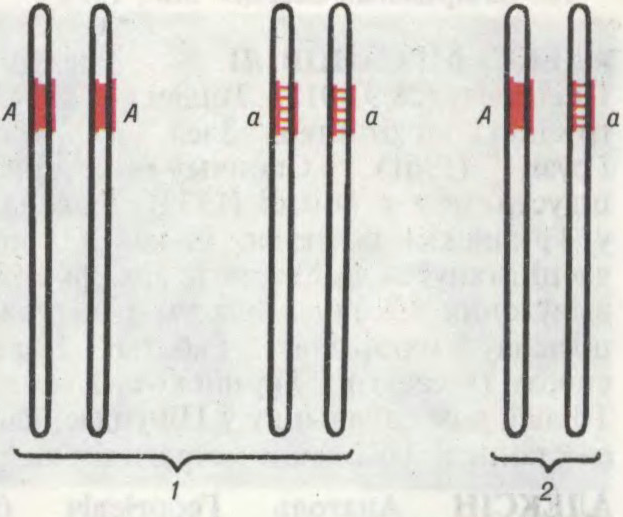
алеламорфы, розныя формы (станы) аднаго і таго ж гена. Размешчаны ў аднолькавых участках (локусах) гамалагічных (парных) храмасом і кантралююць кірункі развіцця адной і той жа прыкметы (напр., белы ці чырвоны колер кветкі). Алелі ўзнікаюць пры любой змене структуры гена ў выніку мутацый або ўнутрыгенных рэкамбінацый (магчымая колькасць алеляў кожнага гена практычна незлічоная). Кожны ген можа знаходзіцца не менш як у двух алельных станах, адзін з якіх звычайна забяспечвае максімальнае развіццё прыкметы — дамінантная алеля, другі прыводзіць да частковай або поўнай страты або змены прыкметы — рэцэсіўная алеля. Ген, які мае некалькі розных станаў, утварае серыю множных алеляў. Наяўнасцю алельных генаў абумоўлены фенатыпічныя адрозненні сярод арганізмаў (гл.Фенатып). Частоты асобных алеляў у генафондзе даюць магчымасць вылічаць генет. змены ў пэўнай папуляцыі і вызначаць частату генатыпаў, што выкарыстоўваюць у селекцыі для прадказання магчымых вынікаў скрыжаванняў.
Схема гома- і гетэразіготнасці па адной пары алеляў: 1 — гомазіготнасць; 2 — гетаразіготнасць (А — дамінантная алель, а — рэцэсіўная алель).
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ІНВЕ́РСІЯў біялогіі,
тып храмасомнай перабудовы (мутацыі), які характарызуецца разрывам і паваротам аднаго з унутр. участкаў храмасомы на 180°. Прыводзіць да змены чаргавання сайтаў у межах гена або генаў у межах храмасомы.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЕТЭРАЗІГО́ТНАСЦЬ (ад гетэра... + зігота),
спадчынная неаднароднасць гібрыднага арганізма, звязаная з наяўнасцю ў яго гамалагічных храмасомах розных форм (алелей) таго ці іншага гена. Узнікае часцей у выніку зліцця разнаякасных паводле геннага або структурнага складу гамет у гетэразіготу, магчыма і праз мутацыі. Звычайна спрыяе высокай жыццяздольнасці арганізмаў і прыстасаванасці іх да зменлівых умоў асяроддзя, таму пашырана ў прыродных папуляцыях, падтрымлівае ў іх пэўны ўзровень генатыпічнай зменлівасці. У эксперыментах гетэразіготнасць атрымліваюць скрыжаваннем паміж сабой гомазігот па розных алелях. Нашчадкі ад такога скрыжавання аказваюцца гетэразіготамі паводле дадзенага гена. Маскіруючае дзеянне дамінантных алелей пры гетэразіготнасці — прычына захавання і пашырэння ў папуляцыі шкодных рэцэсіўных алелей, якія выяўляюць звычайна пры племянной і селекцыйнай рабоце; у мед. генетыцы важная пры вызначэнні спадчынных захворванняў. Гл. таксама Гомазіготнасць.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДУПЛІКА́ЦЫЯ (ад лац. duplicatio падваенне),
разнавіднасць храмасомнай перабудовы, у выніку якой узнікае паўтарэнне ўчастка гена ці храмасомы. Паводле Д. гетэразіготныя арганізмы нясуць 2 дозы дупліцыраваных генаў, гомазіготныя — 4. Адбываецца пры разрыве і абмене ўчасткамі храмасом, а таксама ў выпадку няроўнага кросінговера; можа быць унутрыхрамасомная і міжхрамасомная. Адыгрывае важную ролю пры ўтварэнні новых генаў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДЭЗОКСІРЫБАНУКЛЕА́ЗЫ, ДНКазы,
ферменты класа нуклеаз, якія каталізуюць расшчапленне фосфадыэфірных сувязей у полінуклеатыдных ланцугах ДНК. Шырока прадстаўлены ў клетках жывёл, раслін і мікраарганізмаў. Удзельнічаюць у рэгуляванні распаду і сінтэзу ДНК, аднаўленні структуры яе малекул праз выдаленне пашкоджаных участкаў. Уласцівасць Д. аднаўляць структуры малекул выкарыстоўваюць у геннай інжынерыі пры выдаленні ці ўманціраванні ў малекулу ДНК патрэбнага гена.