ЛЯ́МПА АДВАРО́ТНАЙ ХВА́ЛІ,
электравакуумная
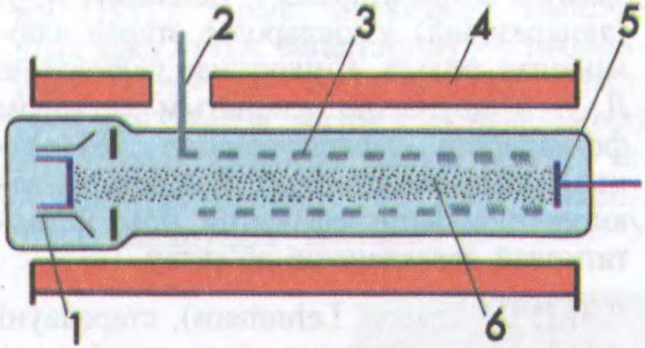
Прынцып работы Л.а.х. вызначыў
А.А.Кураеў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЛЯ́МПА АДВАРО́ТНАЙ ХВА́ЛІ,
электравакуумная
Прынцып работы Л.а.х. вызначыў
А.А.Кураеў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
О́ПЕНГЕЙМЕР ((Oppenheimer) Роберт) (22.4.1904, Нью-Йорк — 18.2.1967),
амерыканскі фізік-тэарэтык, заснавальнік
Тв.:
Літ.:
Рузе М. Роберт Оппенгеймер и атомная бомба:
М.М.Касцюковіч.

Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
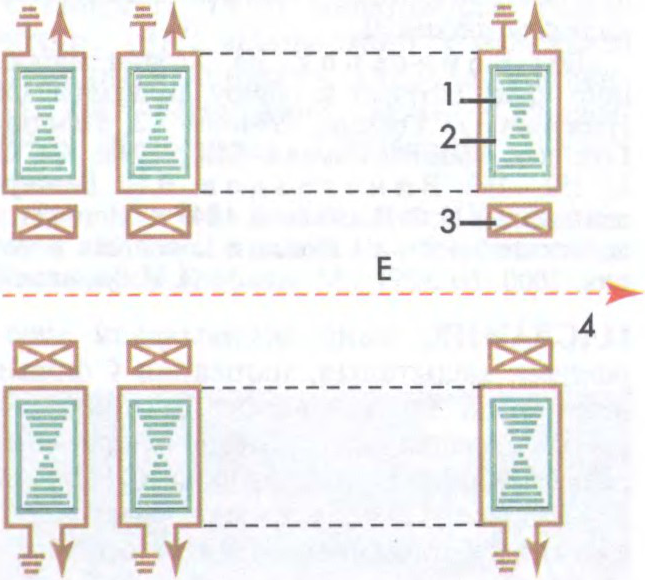
ПАСКАРА́ЛЬНІК З СУСТРЭ́ЧНЫМІ ПУЧКА́МІ,
устаноўка, у якой ажыццяўляюцца сутыкненні сустрэчных пучкоў зараджаных часціц, паскораных
Бываюць цыклічныя (
І.С.Сацункевіч.


Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГО́РЫ,
участкі зямной паверхні, якія ўзняты вышэй за 500
Горы адасоблены ад сумежных раўнін падножжамі ці пераходнай паласой невысокіх (500 — 700
Літ.:
Гвоздецкий Н.А., Голубчиков Ю.Н. Горы.
Леонтьев О.К., Рычагов Г.И. Общая геоморфология. 2 изд.
Л.У.Мар’іна.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АЛІГАФРЭНІ́Я (ад аліга... +
разумовая недаразвітасць, прыроджаная ці набытая ў першыя гады жыцця дзіцяці. Прычыны: паталогія на ўзроўні генаў і храмасом, неспрыяльнае ўздзеянне экзагенна-
Дзеці з
Ф.М.Гайдук.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
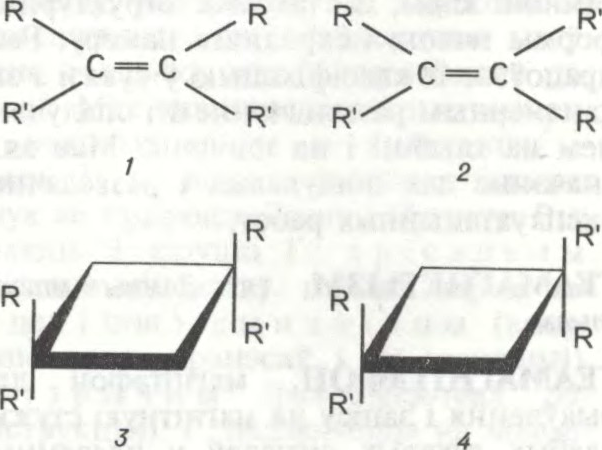
ГЕАМЕТРЫ́ЧНАЯ ІЗАМЕ́РЫЯ,
з’ява існавання малекул рознай прасторавай
Геаметрычная ізаметрыя ўласцівая злучэнням з падвойнымі сувязямі (найчасцей C=C і C=N), вакол якіх немагчыма свабоднае вярчэнне атамаў, і цыклічным злучэнням з малымі (неараматычнымі) цыкламі. Магчыма, калі атам вугляроду пры падвойнай сувязі ці ў цыкле мае неаднолькавыя замяшчальнікі (групоўку атамаў тыпу RR′C = CRR′), якія па-рознаму размешчаны адносна плоскасці падвойнай сувязі (
Літ.:
Потапов В.М. Стереохимия. 2 изд.
М.Р.Пракапчук.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БРЫТА́НСКАЯ ЭНЦЫКЛАПЕ́ДЫЯ,
В.К.Шчэрбін.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
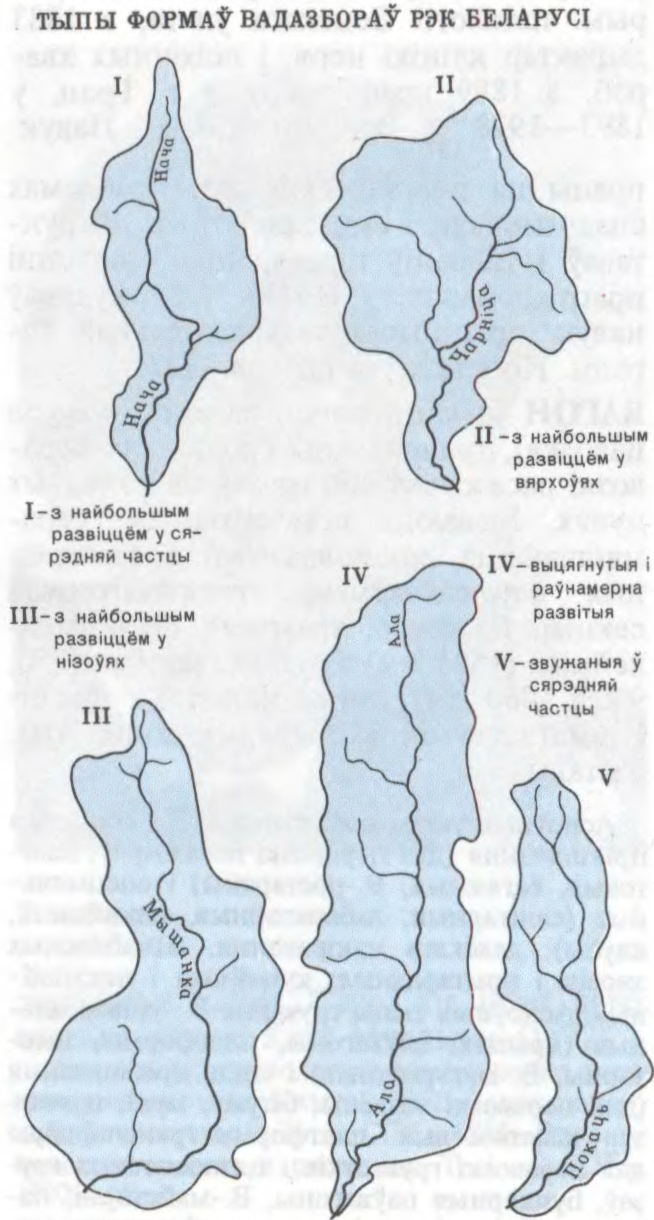
ВАДАЗБО́Р,
1)
2)
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДЭВІЯ́НТНЫЯ ПАВО́ДЗІНЫ,
паводзіны з адхіленнем ад агульнапрынятых у грамадстве прававых і маральна-этычных норм.
Э.С.Дубянецкі.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДЭВІЯ́ЦЫЯ (ад позналац. deviatio адхіленне),
1) адхіленне рухомых цел (карабля, самалёта, снарада, кулі і
2) Адхіленне рухомай сістэмы компаса ад напрамку на
3) Адхіленне частаты ваганняў ад сярэдняга значэння (
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)