парадак двухдольных раслін. Найб. прымітыўныя з існуючых кветкавых раслін 8 сям., 174 роды, каля 3 тыс. відаў. Пашыраны пераважна ў трапічных і субтрапічных паясах. На Беларусі 2 інтрадукаваныя віды з родаў магнолія і лірыядэндран (гл.Цюльпаннае дрэва) сям магноліевых.
Вечназялёныя і лістападныя дрэвы і кусты. Лісце з прылісткамі або без іх. Кветкі спіральныя, спірацыклічныя, часта з доўгай воссю, адзіночныя ці ў суквеццях. Калякветнік просты або дыферэнцыраваны на чашачку і вяночак. Тычынкі пераважна шматлікія, часта стужкападобныя. Лек., дэкар., тэхн., харч.расліны.
Літ.:
Жизнь растений. Т. 5, ч. 1. Цветковые растения. М., 1980.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
НІВЯ́НІК (Leucanthemum),
род кветкавых раслін сям. складанакветных. Каля 20 відаў. Пашыраны ў Еўразіі. На Беларусі 1 від Н. звычайны, або папоўнік (L. vulgare, нар. назвы белая ляхаўка, лугавы рамонак, румянак лясны, пастрэл). Трапляецца каля дарог, на лугах, у хмызняку. Мае шмат садовых форм і сартоў пад агульнай назвай садовы рамонак. Разам з Н. найвялікшым (L. maximum) культывуецца як дэкар. расліна.
Шматгадовыя карэнішчавыя травы выш. да 1 м з прамастойнымі галінастымі сцёбламі. Лісце суцэльнае, радзей перысталопасцевае, чаргаванае. Суквецце — буйны адзіночны кошык. Краявыя кветкі несапраўднаязычковыя, белыя, сярэдзінныя — трубчастыя, жоўтыя. Плод — сямянка. Дэкар.расліны.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЛУБЯНЫ́Я КУЛЬТУ́РЫ,
расліны, якія вырошчваюць для атрымання лубяных валокнаў. Паходзяць з розных сям., растуць ва ўмераных, трапічных і субтрапічных зонах. Адна- і шматгадовыя культуры. На Беларусі культывуюць лён-даўгунец, каноплі. Валакно знаходзіцца ў кары сцябла ў абутылону, джуту, лёну-даўгунцу (гл. ў арт.Лён), канапель, кенафу, кендыру, краталярыі, рамі і інш., у лісці агавы (валакно сізаль), новазеландскага лёну, прадзільнага банана (валакно манільская пянька) і інш. Валокны — сыравіна для тэкст. прам-сці, алей насення прыдатны ў ежу і на тэхн. мэты, з кастрыцы робяць буд. пліты, паперу, тэрмаізаляцыйныя матэрыялы, выкарыстоўваюць на паліва і інш.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
МАШТАКО́Ў (Сяргей Міхайлавіч) (24.10.1905, с. Нікалаеўскі Гарадок Саратаўскага р-на, Расія — 28.7.1970),
бел. вучоны ў галіне фізіялогіі раслін. Д-рбіял.н. (1955), праф. (1956). Скончыў Саратаўскі с.-г.ін-т (1930). З 1948 у АН Беларусі: Ін-це меліярацыі (заг. лабараторыі), з 1956 у Ін-це біялогіі (нам. дырэктара), з 1963 у Ін-це эксперым. батанікі (заг. лабараторыі). Навук. працы па біясінтэзе каўчуку, фізіялогіі збожжавых у сувязі з паляганнем, дзеянні інсектафунгіцыдаў і гербіцыдаў на с.-г.расліны.
Тв.:
Гербициды в борьбе с сорной растительностью. Мн., 1960;
Физиологическое действие некоторых гербицидов на растения. Мн., 1971 (у сааўт.).
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
МНІ́Й (Mnium),
род брыевых імхоў сям. мніевых. Пашыраны ва ўмераным поясе і горных абласцях Паўн. паўшар’я. На Беларусі 4 віды М.: абкружаны (M. marginatum), аднагадовы (M. hornum), зорчаты (M. stellare), няпэўны (M. ambiguum). Трапляюцца на глебе, гнілой драўніне, у вільготных і забалочаных месцах.
Адна- і двухдомныя лістасцябловыя расліныдаўж. 2—7 см. Дзярнінкі шчыльныя або рыхлыя, светла- ці цёмна-зялёныя. Сцябло прамастойнае са стэрыльнымі парасткамі.
Лісце скучанае, падоўжана-эліптычнае або падоўжана-ланцэтнае, завостранае. Спарагоны адзінкавыя, рэдка па 2. Каробачка гарызантальная або паніклая, вечка з дзюбкай, каўпачок клабукападобны.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АМБРО́ЗІЯ (Ambrosia),
род кветкавых раслін сям. астравых. Каля 40 відаў. Пашыраны ў Паўн. і Цэнтр. Амерыцы (у іншых краінах — занесеныя). На Беларусі рэдка трапляюцца амброзія палыналістая (A. artemisiifolia) і трохраздзельная (A. trifida), магчымы знаходкі і інш. відаў, якія актыўна пашыраюцца ў Еўропе і з’яўляюцца каранцінным пустазеллем.
Адна- або шматгадовыя травяністыя расліны, радзей паўкусты з галінастым сцяблом выш. да 2 м. Лісце супраціўнае або чаргаванае: перыстарассечанае. Кветкі малапрыкметныя, у маленькіх аднаполых кошыках, сабраных у коласападобныя суквецці (песцікавыя 2-кветныя). Плод — калючкаватая сямянка (адна расліна можа даваць да 90 000 сямянак). Пылок з кветак выклікае алергічныя захворванні: сянную ліхаманку, бранхіяльную астму і інш.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АПЫЛЕ́ННЕ,
перанос пылку на рыльца песціка (у пакрытанасенных) або на семязавязь (у голанасенных) раслін. Папярэднічае апладненню. Адрозніваюць 2 біял. тыпы апылення: самаапыленне (перанос пылку ў межах адной кветкі — аўтагамія або з інш. кветак той жа расліны — гейтанагамія) і перакрыжаванае (перанос пылку з кветак інш. раслін). Самаапыленне менш пашырана, трапляецца ў некаторых дзікарослых (напр., дзьмухавец, ястрабок) і культурных (ячмень, пшаніца, гарох, фасоля) раслін; у некаторых відаў адбываецца ў нераскрытых кветках (клейстагамія). Перакрыжаванае апыленне больш прагрэсіўнае (аб’ядноўваюцца розныя геномы, у выніку ўтвараецца гетэразіготнае, больш жыццяздольнае патомства). Пылок можа пераносіцца ветрам (анемафілія), вадой (гідрафілія), насякомымі (энтамафілія), птушкамі (арнітафілія) і інш. У селекцыі і раслінаводстве шырока выкарыстоўваюць штучнае апыленне.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
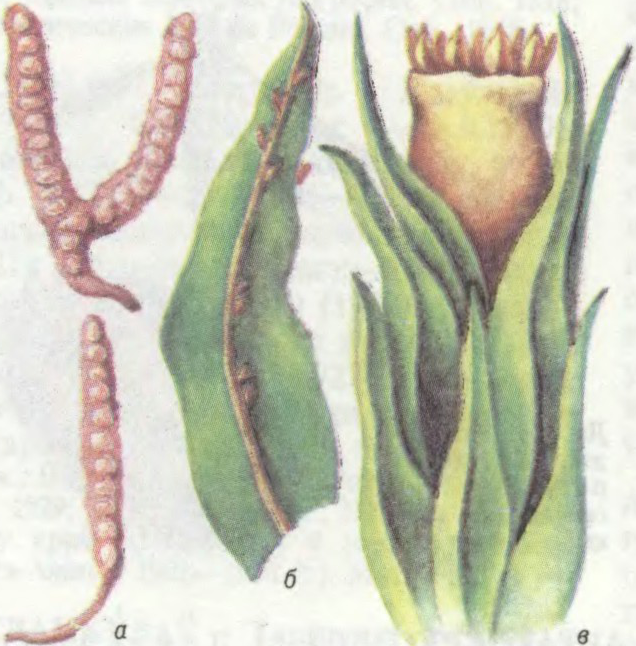
АРТО́ТРЫХУМ (Orthotrichum),
род брыевых імхоў сям. артотрыхавых. Каля 200 відаў. Пашыраны пераважна ва ўмераных шыротах. На Беларусі 15 відаў, з іх артотрыхум прыгожы (O. speciosum) трапляецца часта, артотрыхум Лаеля (O. layelii) занесены ў Чырв. кнігу. Растуць на кары дрэў, камянях, мураваных і бетонных збудаваннях. Ксераморфныя расліны, вытрымліваюць працяглае высыханне.
Падушачкі ад цёмна-зялёных да карычневых. Сцябло 0,5—7 смдаўж., прамастойнае або прыўзнятае, разгалінаванае, густа ўкрытае лісцем. Лісце яйцападобна- або лінейна-ланцэтнае, з простай жылкай. Каробачка на кароткай ножцы, з 8—16 прадаўгаватымі палоскамі, радзей без іх. Вечка конусападобнае з дзюбкай. Перыстом двайны.
Артотрыхум Лаеля: а — вывадковыя цельцы; б — ліст; в — верхняя частка сцябла з каробачкай.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
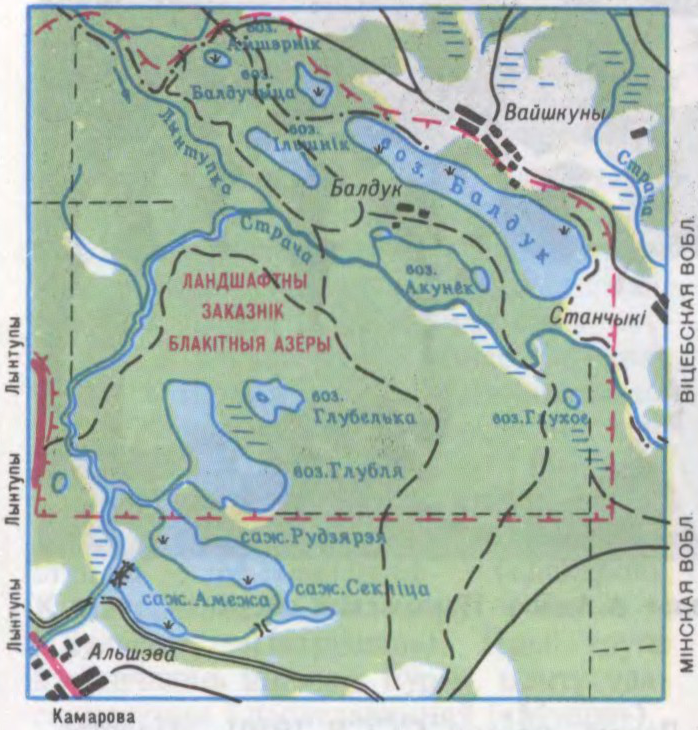
БАЛДУ́ЦКАЯ ГРУ́ПА АЗЁРАЎ,
у Беларусі, на мяжы Пастаўскага р-на Віцебскай вобл., Мядзельскага р-на Мінскай вобласці і Астравецкага р-на Гродзенскай вобл., на ЗБел. Паазер’я, у бас.р. Страча, на тэр. ландшафтнага заказніка Блакітныя азёры. Уключае невял. азёры Балдук, Глубелька, Ячмянец, Мёртвае, Імшарац, Глубля. Агульная пл. 1,5 км².
Утварылася ў час паазерскага зледзянення, размешчана сярод комплексу водна-ледавіковых формаў рэльефу (адносныя выш. 25—30 м), парослага хваёвым і хваёва-яловым лесам. Азёры адметныя чыстай вадой і адсутнасцю антрапагенных экалагічных парушэнняў, у іх растуць рэдкія ахоўныя водныя расліны (хваснік звычайны і меч-трава звычайная). Па азёрах праходзіць турысцкі маршрут.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БАГАМО́ЛЫ (Mantoptera, Mantodea),
атрад драпежных насякомых. Больш за 2000 відаў, пашыраны ў тропіках і субтропіках абодвух паўшар’яў. Найчасцей трапляецца багамол звычайны (Mantis religiosa). Вядомы таксама багамол дрэвавы (Hierodula tenuidentata), рывеціны (Rivetima), багамолы-малышкі (Armene), эмпузы (Empusidae).
Даўж. да 11 см. Цела падоўжанае, галава рухомая трохвугольная, ногі добра развітыя, на пярэдняй пары шыпы, якія ўтвараюць пры складванні моцны хапальны апарат. Ротавыя органы грызучыя. Пярэднія крылы вузкія, заднія складаюцца веерападобна (ёсць бяскрылыя). Яйцы адкладваюць на расліны, камяні; звычайна зімуюць. Драпежнікі, сядзяць у малітоўнай позе (адсюль назва), кормяцца насякомымі, некат. трапічныя віды нападаюць на дробных яшчарак, птушак. Некат. віды занесены ў Чырв. кнігу СССР.