род кветкавых раслін сям. астравых. Больш за 230 відаў. Пашыраны амаль па ўсім зямным шары, асабліва ў Амерыцы. На Беларусі па берагах вадаёмаў, на тарфяніках, у сырых лясах і хмызняках звычайна трапляюцца мясцовыя ваўчкі паніклыя (Bidens cernua) і трохраздзельныя (Bidens tripartita), рэдка — прамяністыя (Bidens radiata), а таксама занесеныя з Паўн. Амерыкі ваўчкі аблісцелыя (Bidens frondosa).
Аднагадовыя травяністыя расліны з прамастойным ці ўзыходным сцяблом. Лісце супраціўнае (верхняе рэдка чаргаванае), простае, цэласнае, 3—5-раздзельнае або рассечанае. Кветкі дробныя, жоўтыя, двухполыя, сабраныя ў гама- або гетэрагамныя кошыкі. Кошыкі адзіночныя або сабраныя ў гронка- ці шчытападобныя агульныя суквецці. Плод — сямянка з 2—4 учэпістымі шчацінкамі. Лек. (мачагонны, патагонны, вітамінны і інш. сродак) і тэхн. (фарбавальныя) расліны; некаторыя віды — пустазелле.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ВЕГЕТАТЫ́ЎНАЕ РАЗМНАЖЭ́ННЕ,
утварэнне новага арганізма з часткі мацярынскага, адзін са спосабаў бясполага размнажэння. У жывёльных арганізмаў ажыццяўляецца пачкаваннем (губкі, кішачнаполасцевыя, некат. чэрві, імшанкі, абалоннікі) або дзяленнем (прасцейшыя, плоскія і кольчатыя чэрві), у ніжэйшых раслін (напр., у водарасцей) — часцей дзяленнем, радзей пачкаваннем (некат. сумчатыя грыбы, напр., дрожджы, некат. базідыяльныя грыбы), у ніжэйшых шматклетачных раслін — распаданнем цела на часткі, здольныя да рэгенерацыі. Вышэйшыя расліны могуць размнажацца карэнішчамі (шматгадовыя травы), чаранкамі (сцябловымі — агрэст, вярба, ружа, таполя; лісцевымі — бягонія, седум; каранёвымі — ажына, вішня, маліна, сліва), адводкамі (яблыня, груша, ягадныя культуры), цыбулінамі (цыбуля, часнок), клубнямі (бульба, тапінамбур) і інш. Вегетатыўнае размнажэнне мае шэраг пераваг перад палавым: лёгкасць размнажэння, хуткае развіццё і ранні пачатак плоданашэння, захаванне ў патомстве прыкмет і ўласцівасцей мацярынскай расліны. Выкарыстоўваецца ў пладаводстве, агародніцтве, лесаводстве.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДРАСЁНАКВЕ́ТНЫЯ (Polygonales),
парадак двухдольных кветкавых раслін. Уключае 1 сям. — драсёнавыя, больш за 30 родаў, каля 800 відаў. Пашыраны амаль па ўсім зямным шары, асабліва ў паўн. умераных паясах. На Беларусі больш за 30 дзікарослых і каля 10 інтрадукаваных відаў з 9 родаў. Найб. вядомыя роды: грэчка, драсён, рэвень, шчаўе. Сярод Д. лек., кармавыя, каштоўныя харч., дубільныя, фарбавальныя і меданосныя расліны, ёсць ядавітыя.
Адна- і шматгадовыя травяністыя расліны, паўкусты, кусты, ліяны, рэдка дрэвы (у тропіках). Сцябло галінастае, вузлаватае, каленчатае. Лісце звычайна чаргаванае, рэдка супраціўнае або кальчаковае, простае, суцэльнае ці лопасцевае, з прылісткамі, якія зрастаюцца каля асновы чаранка ў раструб. Кветкі дробныя, звычайна двухполыя, правільныя, трохчленныя, беспялёсткавыя, сабраныя ў складаныя суквецці. Плод — трохгранны арэшак. Насенне з мучністым эндаспермам.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДРАЦЭ́НА (Dracaena),
род кветкавых раслін сям. драцэнавых. Каля 150 відаў. Пашыраны ў тропіках і субтропіках Усх. паўшар’я. Растуць на горных схілах, у прыбярэжных вільготных лясах. На Беларусі ў аранжарэях і пакоях вырошчваюць Д. пахучую (D. fragrans), стракаталістую, дзярэмскую (D. deremensis), зеленалістую Гукера (D. hookeriana).
Шматгадовазялёныя дрэва- і кустападобныя расліны. Лісце сядзячае, падоўжана-ланцэтападобнае, скурыстае, цвёрдае, зялёнае, шэра-зялёнае або стракатае. Кветкі правільныя, двухполыя, звычайна белыя, крэмаватыя, жаўтаватыя або ружовыя, пахучыя, сабраны на верхавінках у мяцёлкі або пучкі. Плод — ягада. Цвітуць рэдка. Размнажаюць насеннем, верхавінкавымі або сцябловымі чаранкамі. Тэхн. (лаканосныя, фарбавальныя, валакністыя), лек. і дэкар.расліны. Некаторыя дрэвападобныя віды Д., у якіх з надрэзаў кары выцякае хрывава-чырв. смаляністы сок, наз.драконавым дрэвам.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДРЫ́ЖНІК (Briza),
род кветкавых раслін сям. метлюжковых. Каля 10 відаў. Пашыраны ў Еўропе, Пярэдняй Азіі, Паўн. і Паўд. Афрыцы, асабліва ў Міжземнамор’і. На Беларусі паўсюдна на лугах, лясных палянах і ўзлесках трапляецца Д. сярэдні (B. media). Як інтрадукаваная дэкар. расліна вырошчваецца і зрэдку дзічэе Д. вялікі (B. maxima).
Шмат- ці аднагадовыя травяністыя расліны з кароткімі паўзучымі падземнымі парасткамі або без іх. Сцябло прамастойнае, простае або ад асновы галінастае. Лісце сядзячае, лінейнае, пляскатае, шыр. 2—8 мм. Кветкі сабраны ў сплясканыя рамбічныя ці яйцападобныя каласкі, якія ўтвараюць агульнае раскідзістае або сціснутае мяцёлчатае суквецце. Плод — зярняўка. Кармавыя, дэкар. (выкарыстоўваюць для групавых пасадак і на сухія букеты) і лек. (мачагонны, сардэчны, супрацьзапаленчы сродак) расліны.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
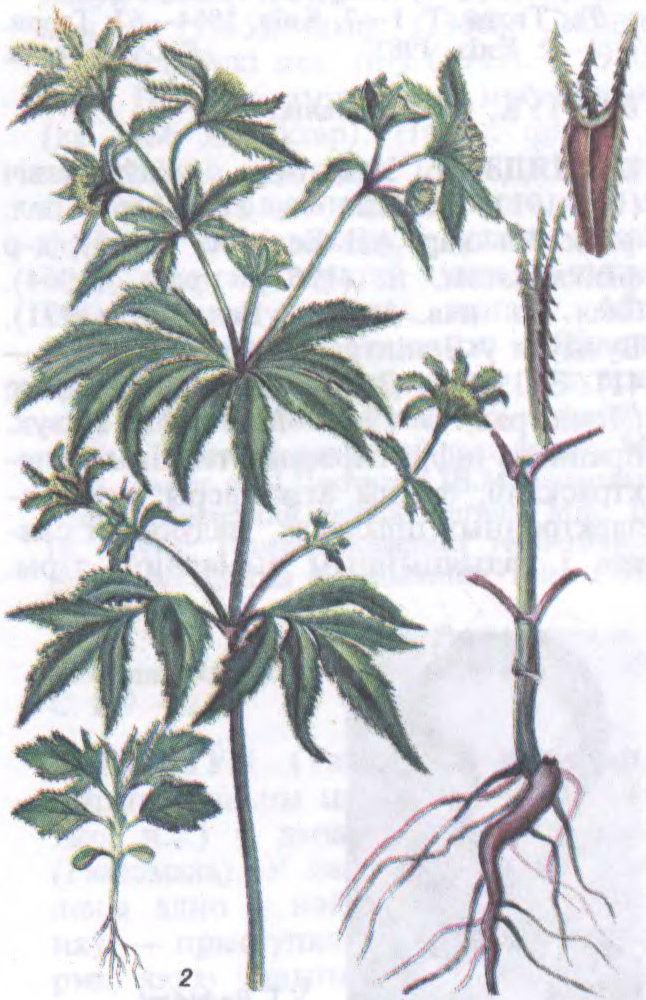
ДУ́ДНІК (Angelica),
род кветкавых раслін сям. сельдэрэевых. Каля 60 відаў. Пашыраны ў Паўн. паўшар’і, асабліва ва Усх. Азіі. На Беларусі 3 дзікарослыя віды: усюды трапляецца Д. лясны (A. sylvestris), зрэдку — лекавы, або дзягіль (A. archangelica), і вельмі рэдка — балотны (A. palustris). Растуць у лясах, хмызняках, на лугах, балотах і інш.Цэнтр.бат. садам Нац.АН Беларусі як дэкар. расліна інтрадукаваны Д. сахалінскі (A. sachalinensis).
Двух- або шматгадовыя травяністыя расліны з кароткім карэнішчам і галінастым, унутры пустым — дудкаватым (адсюль назва) сцяблом выш. да 2 м. Лісце буйное, складанае, перыстарассечанае, з вял. похвамі. Кветкі дробныя, белыя ці ружаватыя, у буйных складаных парасоніках. Плод — віслаплоднік з 10 крылатымі рэбрамі. Лек., кармавыя, эфіраалейныя, меданосныя, вострапрыпраўныя і дэкар.расліны. Плады і карані багатыя кумарынам.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КАЗЯЛЕ́Ц (Ranunculus),
род кветкавых раслін сям. казяльцовых. Больш за 600 відаў. Пашыраны ва ўмераным поясе Паўн. паўшар’я. На Беларусі каля 20 відаў, найб. вядомыя К.: едкі (R. acris), нар. назва курыная слепата, паўзучы (R. repens), пякучы (R. Dammula), залацісты (R. auricomus), шарсцісты (R. lanuginosus), шматкветны (R. polyanthemos), кашубскі (R. cassubicus), даўгалісты (R. lingua), ядавіты (R. sceleratus). Растуць на вільготных лугах, у лясах і інш.
Шмат- і аднагадовыя травяністыя раслінывыш. да 100 см з пучком тонкіх каранёвых валаснікоў ці развітым карэнішчам. Лісце пальчата-раздзельнае або суцэльнае. Кветкі пераважна жоўтыя, адзіночныя ці ў суквеццях. Плод — шарападобны шматарэшак. Многія віды К. атрутныя для буйн. раг. жывёлы і авечак. Лек. і дэкар.расліны.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЖЫ́ЦІК (Lolium),
род кветкавых раслін сям. метлюжковых. Каля 10 відаў. Пашыраны ў Еўропе (пераважна ў Міжземнамор’і), Паўн. Афрыцы і Азіі. На Беларусі як занесеныя, натуралізаваныя і пустазелле трапляюцца Ж. п’янючы (L. temulentum), расстаўлены, або льняны (L. remotum), жорсткі (L. rigidum), шматкветкавы (L. multiflorum) і шматгадовы (L. perenne). Ж. шматкветкавы і шматгадовы пад назвай райграс уведзены ў культуру як кармавыя і газонныя расліны. Некат. віды — пустазелле; ёсць ядавітыя. Растуць каля дарог, на пустках, газонах і інш.
Шмат-, двух- і аднагадовыя травяністыя расліны з прамастойнымі ці ўзыходнымі сцёбламі выш. да 1 м і моцнай валасніковістай каранёвай сістэмай. Лісце вузкалінейнае. Агульнае суквецце — рэдкі лінейны двухрадны колас. Каласкі адзіночныя, сядзячыя, сціснуты з бакоў, з 3—15 кветкамі. Плод — зярняўка.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЗААФА́ГІ [ад заа... + ...фаг(і)],
жывёлы і расліны, кормам для якіх з’яўляюцца інш. жывёлы. Да З. належаць таксама жывёлы, што кормяцца прадстаўнікамі свайго віду (гл.Канібалізм), і паразіты жывёл. Спосаб жыцця З. залежыць ад віду ежы, спосабаў яе здабычы, морфафізіял. і эталагічнай адаптацыі. У актыўных драпежнікаў ёсць органы захопу, умярцвення здабычы, вельмі развіты органы руху і пачуццяў. Стрававальны тракт З. звычайна адносна карацейшы, чым у фітафагаў (арганізмы, якія кормяцца раслінамі). Іх наз.драпежнікамі, хоць у некат. выпадках гэты тэрмін не зусім падыходзіць (напр., лешч з’яўляецца З., аднак драпежнікам яго не называюць). З раслін да З. належаць насякомаедныя расліны (напр., расянка круглалістая, тлушчанка звычайная), з жывёл — ваўкі, лісы, ястрабы, краты і інш.Гл. таксама Некрафагі, Капрафагія, Эўрыфагія, Драпежнікі, Драпежныя звяры, Сокалападобныя.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЗО́РКАЎКА (Stellaria),
род кветкавых раслін сям. гваздзіковых. Каля 120 відаў. Пашыраны амаль па ўсім зямным шары. На Беларусі 10 відаў: З. багнавая (S. alsine), балотная (S. palustris), доўгалістая (S. longifolia), дуброўная (S. nemorum), злакавая (S. graminea), ланцэтападобная (S. holostea), незаўважаная (S. neglecta), сярэдняя, або макрыца (S. media), таўсталістая (S. crassifolia), тупачашачкавая (S. hebecalyx). Растуць у лясах, на лугах, балотах, у хмызняках, на палях, агародах і інш.
Шматгадовыя, радзей двух- і аднагадовыя травяністыя расліны з простым ці галінастым сцяблом выш. да 60 см. Лісце супраціўнае, ад лінейна-ланцэнтнага да яйцападобнага. Кветкі двухполыя, 5-, рэдка 4-членныя, звычайна белыя ў паўпарасонікавых суквеццях. Плод — каробачка. Лек., кармавыя, харч., меданосныя і дэкар.расліны. Многія віды пустазелле, ёсць ядавітыя.