ІНДУ́КЦЫЯ
дынамічнае ўзаемадзеянне нервовых працэсаў узбуджэння і тармажэння. Выражаецца ў тым, што тармажэнне ў групе
А.С.Леанцюк.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ІНДУ́КЦЫЯ
дынамічнае ўзаемадзеянне нервовых працэсаў узбуджэння і тармажэння. Выражаецца ў тым, што тармажэнне ў групе
А.С.Леанцюк.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЛІСТАПА́Д,
фізіялагічны працэс у дрэў, кустоў, радзей траў, звязаны з ападаннем лісця. Выпрацаванае ў працэсе эвалюцыі прыстасаванне да памяншэння паверхні наземных органаў у неспрыяльных умовах. Скарачае страту вільгаці і прадухіляе паломку галін пад цяжарам снегу. Перад Л. у лісці адбываюцца біяхім.,

Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
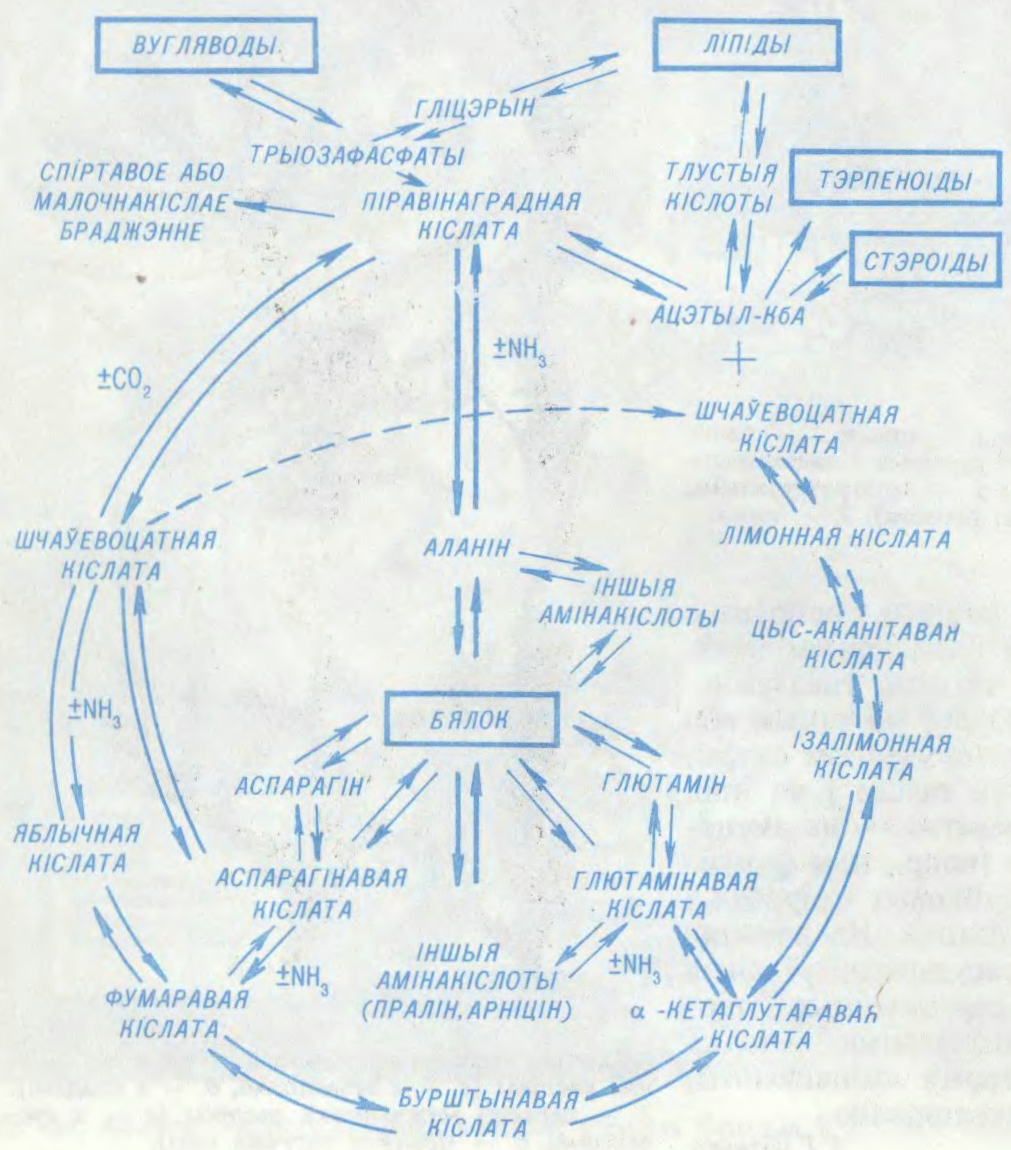
АБМЕ́Н РЭ́ЧЫВАЎ,
сукупнасць
Ператварэнні рэчываў ад моманту іх паступлення ў арганізм да ўтварэння канчатковых прадуктаў распаду складаюць сутнасць
Літ.:
Ленинджер А. Основы биохимии:
Страйер Л. Биохимия:
Я.В.Малашэвіч.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АНАБАЛІ́ЗМ (ад
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ВО́ДНЫ РЭЖЫ́М РАСЛІ́Н,
працэс водаабмену паміж раслінамі і навакольным асяроддзем, неабходны для падтрымання іх жыццядзейнасці; частка агульнага абмену рэчываў. Вызначаецца і ажыццяўляецца ў адпаведнасці з генетычна замацаванымі асаблівасцямі
Л.Г.Емяльянаў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДЫЯБЕ́Т ЦУКРО́ВЫ,
эндакрыннае захворванне, абумоўленае поўным (інсулінзалежны Д.ц.) або адносным (інсуліннезалежны Д.ц.) недахопам у арганізме гармону інсуліну або яго нізкай
Літ.:
Балаболкин М.И. Сахарный диабет.
Справочник по клинической эндокринологии.
Т.В.Мохарт.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ІНСТЫТУ́Т ФОТАБІЯЛО́ГП
Асн. кірункі даследаванняў: малекулярна-мембранная біяхімія і біяфізіка фотабіял. і рэгулятарных працэсаў у жывёльных і раслінных сістэмах (будова, функцыянаванне і біягенез фотасінтэтычнага апарату, фізіялогія і генетыка фотасінтэзу, фотарэцэптарныя працэсы ў раслінных і жывёльных сістэмах, структурная дынаміка і напружаныя метастабільныя станы бялкоў і
Л.Ф.Кабашнікава.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
НЕЙРАЭНДАКРЫНАЛО́ГІЯ (ад нейра... + эндакрыналогія),
раздзел эндакрыналогіі, які вывучае
Зараджэнне Н. звязана з працамі Э.Шарэра (увёў тэрмін «нейрасакрэцыя», 1928) і Б.Шарэра (выявіў сакрэцыю ў
На Беларусі праблемы Н. распрацоўваюцца ў Ін-тах фізіялогіі і біяхіміі
Літ.:
Нейроэндокринология. Ярославль, 1999.
А.А.Холадава.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
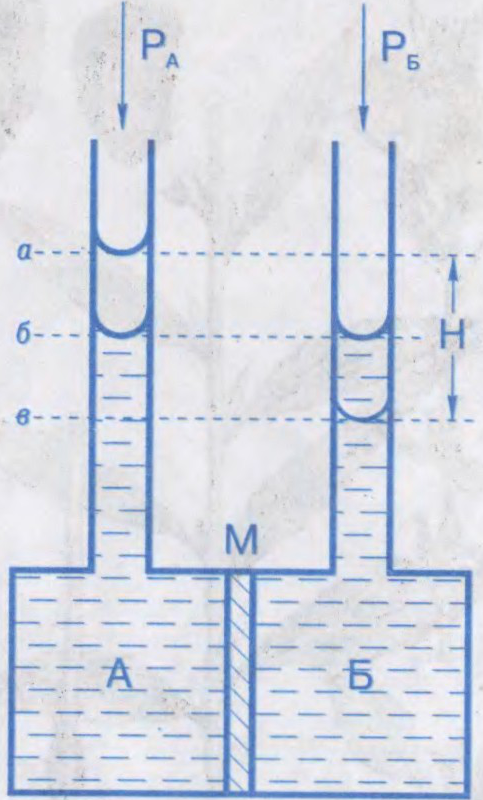
АСМАТЫ́ЧНЫ ЦІСК,
лішкавы гідрастатычны ціск раствору, які перашкаджае дыфузіі растваральніку праз паўпранікальную перагародку; тэрмадынамічны параметр. Характарызуе імкненне раствору да зніжэння канцэнтрацыі пры сутыкненні з чыстым растваральнікам пры сустрэчнай дыфузіі малекул растворанага рэчыва і растваральніку. Абумоўлены змяншэннем хімічнага патэнцыялу растваральніку ў прысутнасці растворанага рэчыва. Роўны лішкаваму вонкаваму ціску, які неабходна прыкласці з боку раствору, каб спыніць осмас. Вымяраецца ў паскалях.
Вымярэнні асматычнага ціску пачаў у 1877
Асматычны ціск адыгрывае важную ролю ў жыццядзейнасці жывых
Літ.:
Курс физической химии.
Пасынский А.Г. Коллоидная химия. 3 изд.
Гриффин Д., Новик
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЕРМАФРАДЫТЫ́ЗМ,
наяўнасць прыкмет мужчынскага і жаночага полу ў аднаго арганізма. Бывае натуральны, уласцівы вышэйшым і ніжэйшым раслінам (аднадомнасць, гоматалізм) і беспазваночным жывёлам (чэрві, малюскі, ракападобныя і
У чалавека сапраўдны гермафрадытызм трапляецца рэдка; характэрна наяўнасць яечніка і яечак або змешанай залозы, часцей мужчынскі набор палавых храмасом (46 XY). Другасныя палавыя прыкметы, як правіла, маюць элементы абодвух полаў. Гермафрадытызм несапраўдны бывае вонкавы і ўнутраны. Пры вонкавым мужчынскім гермафрадытызме ёсць мужчынскія палавыя залозы, а вонкавыя палавыя органы падобныя да жаночых; пры ўнутраным — побач з яечнікамі, недаразвітай прадстацельнай залозай і семявымі пузыркамі могуць быць матка і матачныя трубы. Зрэдку трапляюцца выпадкі жаночага несапраўднага гермафрадытызму, калі сфарміраваны яечнікі, а вонкавыя палавыя органы і другасныя палавыя прыкметы развіваюцца па мужчынскім тыпе. Лячэнне гермафрадытызму: аператыўная змена полу. Дзетанараджэнне немагчыма.
Літ.:
Савченко Н.Е. Гипоспадия и гермафродитизм.
Голубева И.В. Гермафродитизм.
М.Я.Саўчанка.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)