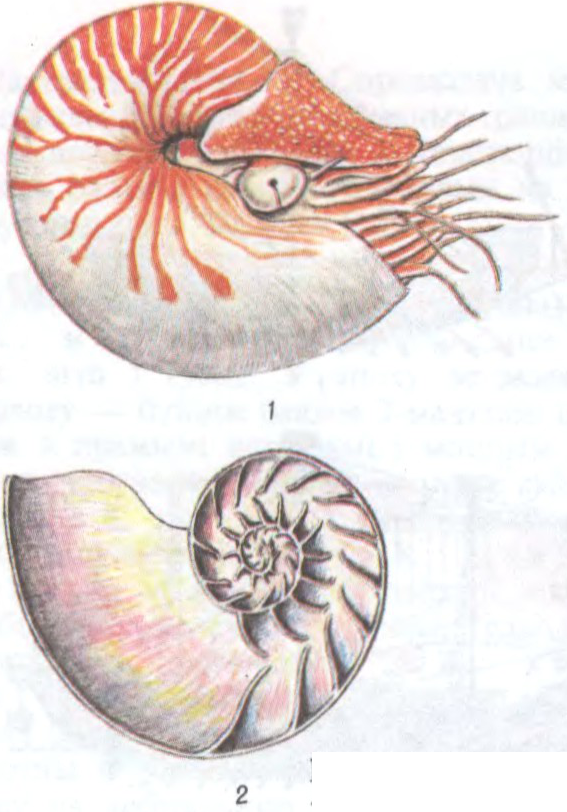
род беспазваночных жывёл кл. галаваногіх малюскаў. 6 відаў. Пашыраны ва ўсх.ч. Індыйскага і зах.ч. Ціхага акіянаў. Жывуць на роўным дне, на глыб. да 750 м.
Дыяметр ракавіны да 30 см, маса да 1,8 кг. Ракавіна спіральная, знутры ўкрыта перламутрам. Падзелена на камеры, якія пранізаны трубкай. Цела жывёлы знаходзіцца ў апошняй, самай вял. камеры, у астатніх — вада ці газ; яны выконваюць ролю гідрастатычнага апарата. Асн. орган пачуццяў — шчупальцы (у самцоў 66, у самак 94), без прысоскаў. Кормяцца ракападобнымі, рыбамі. Ядомыя.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КАРЫЯЛІ́СА СІ́ЛА,
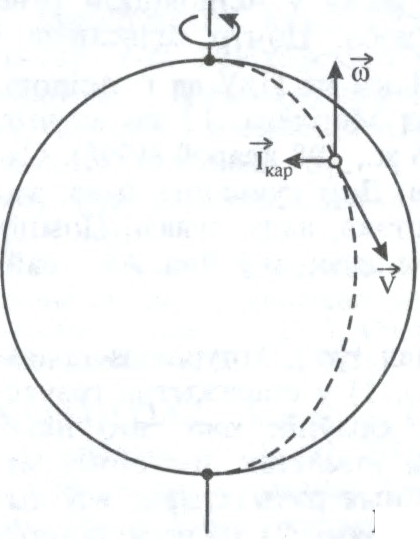
адна з сіл інерцыі, абумоўленая ўплывам вярчальнага руху сістэмы адліку на рух цела адносна гэтай сістэмы. К.с.
, дзе m — маса цела, — Карыяліса паскарэнне. Наяўнасць К.с. вядзе да таго, што цела пры сваім руху адхіляецца ў напрамку, перпендыкулярным да яго адноснай скорасці, або дзейнічае на сувязі механічныя, што перашкаджаюць такому адхіленню. К.с. праяўляецца пры руху масіўных цел (напр., балістычных ракет, цяжкавагавых паяздоў), вял. мас паветра і вады (гл.Бэра закон) і інш. Названа ў гонар Г.Г.Карыяліса.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КРЭ́ЧАТ (Falko rusticolus, або F. gyrfalco),
птушка сям. сакаліных атр. драпежных. Пашыраны на Пн Еўразіі і Паўн. Амерыкі. Селіцца на скалістых узбярэжжах мораў і ў лесатундры. Жыве парамі, устойлівымі многія гады. Усюды рэдкі, сусв. папуляцыя ацэньваецца ў некалькі тысяч пар. Падлягае ахове.
Самы буйны з сапр. сокалаў: даўж. да 60 см, маса самца да 1,5 кг, самкі да 2 кг. Апярэнне шэрае або белае з шэрымі меткамі, верх цямнейшы. Корміцца пераважна птушкамі, грызунамі. Гнёзды на скалах, абрывістых берагах ці высокіх дрэвах. Нясе 3—5 яец. Лоўчая птушка для сакалінага палявання.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КУРАПА́ТКІ,
птушкі сям. цецеруковых і фазанавых атр. курападобных. Пашыраны ўсюды, ад тундры да трапічных лясоў і высакагор’яў. На Беларусі 2 віды: К. белая (Lagopus lagopus) і К. шэрая (Perdix perdix). Жывуць на палях, балотах, лугах, у драбналессі. К. белая занесена ў Чырв. кнігу Беларусі.
Даўж. да 46 см, маса да 900 г. Афарбоўка шэра-бурая, рыжа-бурая, у К. белай зімой белая. Кормяцца парасткамі, пупышкамі, насеннем траў, зернем, насякомымі (у т. л. каларадскімі жукамі). Нясуць да 24 яец. Аб’ект палявання. Гл. таксама Белыя курапаткі.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЛАГО́ЙСКАЯ АСТРАБЛЕ́МА,
старажытны метэарытны кратэр паблізу г. Лагойск у вярхоўі р. Гайна. Пахаваны пад антрапагенавымі адкладамі. Адкрыты ў 1975 пры бурэнні свідравіны ў раёне в. Кузевічы як структура з анамальнай геал. будовай. Дыяметр Л.а. каля 15 км, глыбіня каля 500 м. Складзена з т. зв. брэкчыі — абломкаў, глыб, друзу парод рознага ўзросту; частка іх дэфармаваная, сплаўленая; уся маса кавалкаў сцэментаваная ў больш позні час. Метэарытнае паходжанне Л.а. абгрунтавалі бел. вучоныя А.С.Махнач, М.В.Вераценнікаў, Г.І.Ількевіч.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДУ́ПЕЛЬ (Gallinago media),
птушка сям. сяўцовых атр. сеўцападобных. Пашыраны ў Еўропе і Паўн. Азіі на балотах і заліўных лугах. Пералётны. На Беларусі звычайны, нар. назва дубельт.
Даўж. цела да 35 см, маса да 230 г. Апярэнне стракатае, рыжавата-бурае. Падобны на бакаса, але на баках грудзей і брушка папярочныя стракаціны, больш кароткая дзюба (да 7,4 см). Самцы такуюць вял. групамі ўначы. Гнёзды на купінах. Нясе 4 яйцы. Птушаняты вывадкавыя. Корміцца глебавымі беспазваночнымі, насякомымі і іх лічынкамі, карэньчыкамі балотных раслін. Зімуе ў Паўд.-Зах. і Паўд. Афрыцы. Аб’ект палявання.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ДЫСТРАФІ́Я АЛІМЕНТА́РНАЯ, хвароба галодная,
хвароба, што развіваецца пры адсутнасці ці недастатковым паступленні харч. рэчываў у арганізм. Адрозніваюць 3 стадыі Д.а. На 1 -й стадыі хваробы зніжаецца маса, павышаюцца апетыт, смага, лёгкі ацёк, на 2-й — значныя пахудзенне і ацёкі, мускульная слабасць, цяжкія запоры, паніжэнне т-ры цела, на 3-й стадыі знікае тлушчавая клятчатка, атрафіруюцца мускулы, слабасць, ацёкі ўсяго цела, парушэнне псіхікі. Ускладненні: падае артэрыяльны ціск, анемія, лейкацытапенія, дыспепсія, полінеўрыты, паркінсанізм, зніжэнне памяці, інтэлекту і інш. Лячэнне: паўнацэннае (у невял. дозах) і частае харчаванне, глюкоза, бялковыя прэпараты, пераліванне крыві, саграванне.
клас бялкоў, прастэтычныя групы якіх складаюцца з негімінавага жалеза, комплексна звязанага з неарган. або арган. (у складзе цыстэіну) серай з утварэннем аднаго ці некалькіх Fe—S-кластэраў. Кластэры здольныя аддаваць ці прымаць электроны (але не вадарод) з абарачальнай зменай валентнасці атамаў жалеза (Fe3+ ⇄ Fe2+). Дзякуючы гэтаму Ж. ў жывых арганізмах выконваюць функцыю акісляльна-аднаўленчых кампанентаў у тэрмінальных ланцугах пераносу электронаў паміж флавапратэідамі і цытахромамі (гл.Дыханне). Малекулярная маса Ж. ад 6000 (ферыдаксіны бактэрый) да 12 000 (ферыдаксіны хларапластаў) і больш (ксантынаксідаза).
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЖАРЛЯ́НКА ЧЫРВАНАБРУ́ХАЯ (Bombina bombina),
земнаводнае сям. круглаязычных атр. бясхвостых земнаводных. Пашырана ў Цэнтр. і Усх. Еўропе. Жыве ў плыткіх і зарослых вадаёмах з глеістым дном, на балотах, у лужынах і інш. На Беларусі трапляецца пераважна ў паўд., паўд.-зах. і цэнтр. раёнах.
Даўж. цела 3,5—4,5 см (зрэдку да 6), маса 3—10 г. Спіна светла-шэрая, бураватая з цёмнымі плямамі, бруха ярка-аранжавае або чырвонае з сінявата-чорнымі плямамі. Скура з бародаўкамі, што выдзяляюць едкі сакрэт Актыўная днём і вечарам. Карысная. Корміцца воднымі беспазваночнымі.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЛА́РМАРА ПРЭЦЭ́СІЯ,
прэцэсія аднолькавых зараджаных часціц (напр., электронаў у атаме) у пастаянным аднародным слабым магн. полі. Апісана Дж.Лармарам у 1895. Пакладзена ў аснову класічнай тэорыі дыямагнетызму, тлумачэння Зеемана з’явы і магн. вярчэння плоскасці палярызацыі.
Абумоўлена ўздзеяннем на часціцы Лорэнца сілы; падобная на прэцэсію восі гіраскопа пад уздзеяннем сілы, якая імкнецца павярнуць вось вярчэння. Л.п. адбываецца вакол напрамку магн. поля з вуглавой скорасцю ωL (лармараўская частата), якая вызначаецца. па формуле ωL = qB/(2m), дзе q — зарад, m — маса і q/m — удзельны зарад часціцы, B — індукцыя магн. поля.