1) у жывёл — група асобін, што маюць падобныя морфаэкалагічныя прыстасаванні для пражывання ў аднолькавым асяроддзі. Характэрна змена Ж.ф. у антагенезе для відаў, развіццё якіх ідзе з метамарфозам (ад лічынкі да дарослага насякомага). У аснове класіфікацыі жывёл — асаблівасці размнажэння, эколага-марфал. прыстасаванні, спосабы перамяшчэння, здабычы ежы і яе характар, аднесенасць да ландшафту, ступень актыўнасці і інш. 2) Ж.ф. ў раслін — вонкавы выгляд (габітус), які адлюстроўвае іх прыстасаванасць да ўмоў асяроддзя; адзінка экалагічнай класіфікацыі. Складаюцца ў выніку натуральнага адбору ў пэўных умовах асяроддзя. Ж.ф. груп раслін адлюстроўваюць іх прыстасаванасць да рассялення і замацавання на тэрыторыі, да найб. поўнага выкарыстання ўсяго комплексу ўмоў месцазнаходжання. Змяняецца ў антагенезе (напр., аднагадовыя сеянцы елкі або дуба яшчэ не маюць формы дрэва), таму пад Ж.ф. класіфікацыйнай адзінкай лічыцца сукупнасць дарослых асобін. Адзін і той жа від раслін у розных умовах можа мець розныя Ж.ф. (дуб, елка, ядловец і інш. ў лясной зоне або лясным поясе гор — высакастволыя дрэвы, а на паўн. і вышыннай межах арэала — кусты або сланікі). У класіфікацыю можа ўваходзіць група раслін з падобнымі прыстасавальнымі структурамі, не абавязкова звязаных роднасцю (напр., кактусы і некат. малачаі ўтвараюць формы сцябловых сукулентаў). Тэрмін Ж.ф. прапанаваў дацкі батанік І.Э.Вармінг (1884). Пры гэтым ён разумеў форму, у якой вегетатыўнае цела расліны знаходзіцца ў гармоніі з навакольным асяроддзем на працягу ўсяго жыцця, ад насення да адмірання. Найб. пашырана класіфікацыя Ж.ф., прапанаваная дацкім батанікам К.Раўнкіерам (1905, 1907), які вылучае 5 асн. Ж.ф. (гл.Сістэма Раўнкіера). Змяненні Ж.ф. выкарыстоўваюцца для работ па інтрадукцыі раслін.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БАЛОТАЗНА́ЎСТВА,
вучэнне пра балоты, іх паходжанне, развіццё, будову і гасп. выкарыстанне. Мае батаніка-геаграфічны (фітацэналагічны) і тарфазнаўчы кірункі.
Балотазнаўства ўзнікла ў канцы 17 — пач. 18 ст. ў Зах. Еўропе. Інтэнсіўна развівалася ў канцы 19 ст. ў сувязі з ростам меліярац. работ. Заснавальнік балотазнаўства ў Расіі — М.В.Ламаносаў, які даследаваў прыроду торфу. На Беларусі вывучэнне балотаў пачалося з Палесся (М.Бутрымовіч, 1772). Пазней была арганізавана Заходняя экспедыцыя па асушэнні балотаў пад кіраўніцтвам І.І.Жылінскага (1873—98), у якой прынялі ўдзел глебазнавец В.В.Дакучаеў і батанік Г.І.Танфільеў, што апублікаваў першую ў Расіі класіфікацыю балотаў і апісаў расліннасць балотаў Палесся (1899). У 1911 адкрыта Мінская балотная доследная станцыя. Беларусь стала цэнтрам вывучэння і практычнага выкарыстання балотаў Расіі.
У пач. 20 ст.вял. ўклад у развіццё балотазнаўства зрабілі бел. вучоныя У.М.Сукачоў, А.Ф.Флёраў, В.Р.Вільямс, У.С.Дактуроўскі і інш. У 1925—30 работы па акультурванні балотаў вялі А.Т.Кірсанаў, М.Ф.Лебядзевіч, І.С.Лупіновіч, С.Г.Скарапанаў, іх прадаўжалі А.П.Підоплічка, М.М.Бамбалаў, П.І.Бялькевіч, І.І.Ліштван, У.Я.Ракоўскі, А.В.Цішковіч і інш. Вывучаюць балоты ў НДІ меліярацыі і лугаводства, глебазнаўства і аграхіміі, Ін-це праблем выкарыстання прыродных рэсурсаў і экалогіі АН Беларусі і інш.н.-д. установах. Распрацавана карта тарфяных радовішчаў, абгрунтаваны сыравінныя базы бітумінозных тарфоў і паказаны ўмовы іх залягання. Даследаваны геагр. і экалагічныя асаблівасці развіцця некаторых сфагнавых комплексаў на вярховых балотах. Дадзены асн.генет. схемы развіцця тарфянікаў на тэр. рэспублікі. Вывучаюцца генезіс, глебаўтваральныя працэсы і раслінныя асацыяцыі балотаў, метады рэгулявання воднага рэжыму, праблемы, звязаныя з с.-г. выкарыстаннем асушаных балотаў, прамысл. асваеннем тарфяных радовішчаў і інш. На аснове даследаванняў па балотазнаўстве складаюцца праекты меліярацыі, выкарыстання торфу ў нар. гаспадарцы.
Літ.:
Підоплічка А.П., Смаляк Л.П. Даследаванні балотнай расліннасці Беларусі // Навука БССР за 50 год. Мн., 1968;
Юркевич И.Д., Голод Д.С., Адерихо В.С. Растительность Белоруссии, ее картографирование, охрана и использование. Мн., 1979.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АСМАТЫ́ЧНЫ ЦІСК, дыфузны ціск,
лішкавы гідрастатычны ціск раствору, які перашкаджае дыфузіі растваральніку праз паўпранікальную перагародку; тэрмадынамічны параметр. Характарызуе імкненне раствору да зніжэння канцэнтрацыі пры сутыкненні з чыстым растваральнікам пры сустрэчнай дыфузіі малекул растворанага рэчыва і растваральніку. Абумоўлены змяншэннем хімічнага патэнцыялу растваральніку ў прысутнасці растворанага рэчыва. Роўны лішкаваму вонкаваму ціску, які неабходна прыкласці з боку раствору, каб спыніць осмас. Вымяраецца ў паскалях.
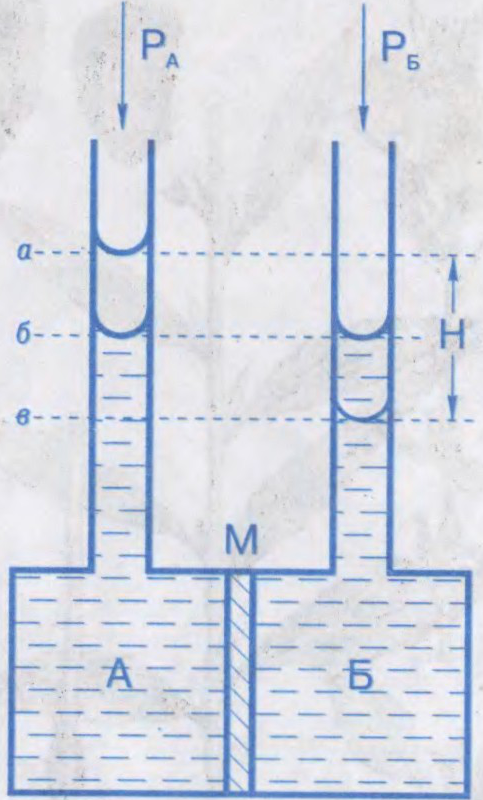
Вымярэнні асматычнага ціску пачаў у 1877 ням.батанік В.Пфефер у растворы трысняговага цукру. Па яго даных галандскі хімік Я.Х.Вант-Гоф устанавіў у 1887, што залежнасць асматычнага ціску ад канцэнтрацыі цукру па форме супадае з Бойля-Марыёта законам для ідэальных газаў. Асматычны ціск вымяраюць з дапамогай асмометраў. Статычны метад вымярэння асматычнага ціску заснаваны на вызначэнні лішкавага гідрастатычнага ціску па вышыні слупка вадкасці H пасля ўстанаўлення стану раўнавагі пры роўнасці вонкавых ціскаў PА і PБ; дынамічны метад зводзіцца да вымярэння скорасці V усмоктвання і выціскання растваральніку з асматычнай ячэйкі пры розных значэннях лішкавага ціску P = PА – PБ з наступнай інтэрпаляцыяй атрыманых даных да V=0 пры лішкавым ціску Δp, роўным асматычнаму ціску. Па велічыні асматычнага ціску распазнаюць: ізатанічныя, або ізаасматычныя, растворы, якія маюць аднолькавы асматычны ціск (незалежна ад саставу), гіпертанічныя з больш высокім Асматычным ціскам і гіпатанічныя растворы з больш нізкім асматычным ціскам.
Асматычны ціск адыгрывае важную ролю ў жыццядзейнасці жывых клетак і арганізмаў. У клетках і біял. вадкасцях ён залежыць ад канцэнтрацыі раствораных у іх рэчываў. Па велічыні асматычнага ціску вадкасцяў унутр. асяроддзя арганізма (кроў, гемалімфа і інш.) водныя арганізмы падзяляюцца на гіпер-, гіпа- і ізаасматычныя. Сярэдняя велічыня і дыяпазон асматычнага ціску ў розных арганізмаў розныя і залежаць ад віду і ўзросту арганізма, тыпу клетак і асматычнага ціску навакольнага асяроддзя (напр., асматычны ціск клетачнага соку наземных органаў балотных раслін 0,2—1,6 МПа, у стэпавых 0,8—0,4, у дажджавых чарвякоў 0,36—0,48, у прэснаводных рыб 0,6—0,66, у акіянічных касцістых рыб 0,78—0,85, акулавых 2,2—2,3, млекакормячых 0,66—0,8 МПа). У гіперасматычных арганізмаў (прэснаводныя жывёлы, некаторыя марскія храстковыя рыбы — акулы, скаты; усе расліны) унутр. Асматычны ціск перавышае асматычны ціск навакольнага асяроддзя, таму іоны могуць актыўна паглынацца арганізмам і ўтрымлівацца ў ім, а вада паступае праз біял. мембраны пасіўна, у адпаведнасці з асматычным градыентам. У гіпаасматычных жывёл (касцістыя рыбы, некаторыя марскія паўзуны, птушкі) асматычны ціск крыві меншы за асматычны ціск навакольнага асяроддзя. Адноснае пастаянства Асматычнага ціску забяспечваецца водна-салявым абменам праз осмарэгулявальныя органы (гл. ў арт.Осмарэгуляцыя).
Літ.:
Курс физической химии. Т. 1—2. 2 изд. М., 1970—73;
Пасынский А.Г. Коллоидная химия. 3 изд. М., 1968;
Гриффин Д., Новик Эл. Живой организм: Пер. с англ.М., 1973.
Вымярэнне асматычнага ціску асмометрам: А — камера для раствору; Б — камера для растваральніка; М — мембрана. Узроўні вадкасці ў трубках пры асматычнай раўнавазе: а, в — пры роўнасці вонкавых ціскаў PА = PБ; б — пры роўнасці рознасці PА−PБ асматычнага ціску.