упарадкаванасць мікраструктуры рэчыва на міжатамных (блізкі парадак) ці на неабмежавана вял. адлегласцях (далёкі парадак). Бывае каардынацыйны — у размяшчэнні часціц рэчыва (існуе ў крышталях; у вадкасцях і аморфных цвёрдых целах — толькі блізкі парадак); арыентацыйны — у арыентацыі часціц (у вадкіх крышталях; у вадкасцях з несіметрычных малекул — блізкі парадак); магнітны — у арыентацыі магн. момантаў (у ферамагнетыках, ферымагнетыках, антыферамагнетыках); у арыентацыі эл. дыпольных момантаў (у сегнетаэлектрыках). Існаванне Д. і б.п. абумоўлена ўзаемадзеяннем паміж часціцамі; асн. прыкмета — сіметрыя крышталёў. Ступень упарадкавання можна мяняць цеплавой, мех. і інш. апрацоўкамі, пры гэтым мяняюцца фіз. ўласцівасці рэчыва.
Літ.:
Уайт Р., Джебелл Т. Дальний порядок в твердых телах: Пер. с англ.М., 1982.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ВАВІ́ЛАВА ЗАКО́Н,
вызначае залежнасць квантавага выхаду фоталюмінесцэнцыі ад даўжыні хвалі ўзбуджальнага святла. Адкрыты С.І.Вавілавым (1924), ляжыць у аснове тэорыі люмінесцэнцыі. Паводле Вавілава закона квантавы выхад фоталюмінесцэнцыі пастаянны ў шырокім інтэрвале даўжынь хваляў узбуджальнага святла і рэзка памяншаецца пры даўжынях хваляў, большых за тую, пры якой назіраецца максімум інтэнсіўнасці спектра люмінесцэнцыі.
Пастаянства квантавага выхаду абумоўлена квантавай прыродай святла, хуткім пераразмеркаваннем энергіі ва ўзбуджаным электронным стане па вагальных і інш. падузроўнях і перадачай энергіі навакольнаму асяроддзю. Вавілава закон выконваецца ў вадкіх і цвёрдых растворах; для складаных малекул у газавай фазе не мае месца, што тлумачыцца захаваннем лішку вагальнай энергіі, набытай у акце паглынання святла. Вавілава закон атрымаў тэарэт. тлумачэнне ў працах бел. вучоных Б.І.Сцяпанава і М.К.Барысевіча.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ВАДАРО́ДНАЯ СУ́ВЯЗЬ,
від трохцэнтравай хімічнай сувязі тыпу A—Hδ+... Bδ-, якая ўзнікае, калі атам вадароду H адначасова злучаны з двума электраадмоўнымі атамамі A і B. З атамам A (вуглярод, азот, кісларод, сера) вадарод злучаны моцнай кавалентнай сувяззю (A—Hδ+). З атамам B (фтор, кісларод, азот, радзей хлор, сера), які мае непадзельную пару электронаў, утварае дадатковую вадародную сувязь (абазначаецца кропкамі). Вадародная сувязь на парадак слабейшая за кавалентную сувязь.
Атамы A і B могуць належаць адной (унутрымалекулярная вадародная сувязь) і розным малекулам (міжмалекулярная вадародная сувязь). Выклікае асацыяцыю аднолькавых (вада, кіслоты, спірты) ці розных малекул у асацыяты і комплексы, уплывае на крышталізацыю, растварэнне, вызначае структуру бялкоў, нуклеінавых кіслот і інш. біялагічна важных злучэнняў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЛІКО́ЛІЗ
(ад грэч. glykys салодкі + ..ліз),
адзін з цэнтральных шляхоў расшчаплення глюкозы з цукроў ці поліцукроў у жывёльных, раслінных і многіх бактэрыяльных клетках; ферментатыўны анаэробны працэс негідралітычнага распаду вугляводаў (пераважна глюкозы) да малочнай к-ты. Адыгрывае значную ролю ў абмене рэчываў жывых арганізмаў, забяспечвае клеткі энергіяй ва ўмовах недахопу кіслароду, а ў аэробных умовах з’яўляецца стадыяй, якая папярэднічае дыханню. У гліколізе адзначаюць 3 этапы (гл. схему). На 1-м этапе (рэакцыі 1—4) малекула глюкозы пераўтвараецца ў 2 малекулы гліцэральдэгід-3-фасфату (з выкарыстаннем 2 фасфатных груп і энергіі, якая выдзяляецца пры гідролізе адэназінтрыфасфату — АТФ). На 2-м этапе (рэакцыі 5, 6) альдэгідная група кожнай з 2 малекул гліцэральдэгід-3-фасфату акісляецца да карбаксільнай, а энергія, якая пры гэтым выдзяляецца, ідзе на сінтэз АТФ з адэназіндыфасфату (АДФ) і неарганічнага фасфату, адначасова адбываецца аднаўленне нікацінамідадэніндынуклеатыду (НАД) да нікацінамідадэніндынуклеатыдфасфату (НАДФ). На 3-м этапе (рэакцыі 7—9) 2 малекулы фасфату, якія далучыліся да цукру на 1-м этапе, пераносяцца назад на АДФ, у выніку чаго ўтвараецца АТФ і кампенсуюцца затраты АТФ на 1-м этапе. Сумарны выхад энергіі пры гліколізе зводзіцца да сінтэзу 2 малекул АТФ (на 1 малекулу глюкозы), якія ўтварыліся ў рэакцыях 5 і 6. У большасці клетак жывёл піруват, які ўтвараецца пры гліколізе, поўнасцю акісляецца ў мітахондрыях да CO2 і H2O. У анаэробных арганізмаў і тканках (шкілетныя мышцы) гідроліз — асн. крыніца клетачнага АТФ. Малекулы пірувату, застаючыся ў цытазоле, могуць пераўтварацца ў залежнасці ад віду арганізма ў лактат (у мышцах, некат. мікраарганізмах) або ў этанол + CO2 (у дражджах, гл.Браджэнне).
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БІЯЛЮМІНЕСЦЭ́НЦЫЯ
(ад бія... + люмінесцэнцыя),
свячэнне жывых арганізмаў, абумоўленае біяхім. працэсамі; від хемалюмінесцэнцыі. Уласціва некаторым бактэрыям, ніжэйшым раслінам, грыбам, насякомым (жукі-светлякі і інш.), беспазваночным, рыбам. Вельмі пашырана сярод марскіх жывёл.
У большасці выпадкаў біялюмінісцэнцыя ўзнікае ў выніку ферментатыўнага акіслення асобных рэчываў — люцыферынаў. Частка малекул люцыферынаў за кошт вызваленай пры гэтым хім. энергіі пераходзіць ва ўзбуджаны стан, пры вяртанні ў асн. стан яны выпраменьваюць святло. Біялюмінісцэнцыя выкарыстоўваецца для асвятлення і прынады здабычы (напр., у глыбакаводных рыб), для перасцярогі, адпужвання або адцягвання ўвагі драпежнікаў, у якасці сігналу для сустрэчы самцоў і самак у шлюбны перыяд (розныя віды светлякоў).
Літ.:
Мак-Элрой У.Д., Зелигер Г.Г. Происхождение и развитие биолюминесценции // Горизонты биохимии: Пер. с англ. М., 1964;
Тарусов Б.Н. Сверхслабое свечение живых организмов. М., 1972.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГІПЕРГУ́К,
пругкія хвалі з частатой 109—1013Гц. Па фізічнай прыродзе не адрозніваецца ад ультрагуку (2·104—109Гц). Існуе гіпергук прыродны (цеплавыя ваганні крышталічнай рашоткі) і штучны (генерыруецца пры дапамозе спец. выпрамяняльнікаў; гл.П’езаэлектрычнасць, Магнітастрыкцыя).
Пругкія хвалі распаўсюджваюцца ў асяроддзі, калі іх даўжыні большыя за даўжыню свабоднага прабегу малекул у газах ці міжатамных адлегласцей у вадкіх і цвёрдых целах. Таму ў газах, у т. л. ў паветры, пры нармальных умовах гіпергук не распаўсюджваецца, у вадкасцях хутка затухае; параўнальна добрыя праваднікі гіпергуку — монакрышталі пры нізкіх т-рах. Гіпергук выкарыстоўваюць для даследавання стану рэчыва, асабліва ў фізіцы цвёрдага цела, для стварэння акустычных ліній затрымкі ў ЗВЧ дыяпазоне і інш. прылад акустаэлектронікі і акустаоптыкі.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЛАНЦУГО́ВЫЯ ХІМІ́ЧНЫЯ РЭА́КЦЫІ,
хімічныя рэакцыі, у якіх узнікненне прамежкавай актыўнай часціцы (атама, свабоднага радыкала, іона) і яе рэгенерацыя (узнаўленне) у кожным элементарным акце рэакцыі выклікаюць вялікую колькасць (ланцуг) пераўтварэнняў зыходных рэчываў у прадукты рэакцыі. Тыповыя Л.х.р. — тэрмічны крэкінг, піроліз, акісленне, радыкальная полімерызацыя, галагеніраванне.
Л.х.р. складаецца з некалькіх элементарных стадый: зараджэнне ланцуга (ініцыіраванне) — утварэнне актыўных часціц (АЧ), якое адбываецца, напр., у выніку дысацыяцыімалекул; працяг ланцуга — узаемадзеянне АЧ з зыходнымі рэчывамі, якое прыводзіць да ўтварэння малекул прадукту рэакцыі і новых АЧ; абрыў ланцуга («знікненне» АЧ) у выніку рэкамбінацыі свабодных радыкалаў, узаемадзеяння АЧ са сценкамі пасудзін ці з інгібітарамі. Адрозніваюць Л.х.р. неразгалінаваныя і разгалінаваныя. У неразгалінаваных на кожную АЧ, якая расходуецца пры працягу ланцуга, узнікае адна актыўная часціца, напр., рэакцыя хларыравання вадароду. Пры ініцыіраванні ўтвараецца свабодны атам хлору (Cl), які ўзаемадзейнічае з малекулай вадароду H2(Cl+H2 → HCl+H) з утварэннем малекулы хлорыстага вадароду HCl і свабоднага атама вадароду, які ўзаемадзейнічае з малекулай Cl2(H+Cl2 → HCl+Cl), і г.д. У разгалінаваных Л.х.р. на адну АЧ, расходаваную пры працягу ланцуга, узнікае некалькі АЧ (2 і больш), адна з іх працягвае першы ланцуг, а інш. пачынаюць новыя, што прыводзіць да разгалінавання. Калі скорасць разгалінавання меншая за скорасць знікнення АЧ, разгалінаваная Л.х.р. адбываецца ў стацыянарным рэжыме. Пры нестацыянарным рэжыме (скорасць знікнення АЧ меншая за скорасць разгалінавання) агульная скорасць ланцуговага працэсу імкліва ўзрастае. Пераход да нестацыянарнага рэжыму адбываецца скачком і разглядаецца як самазагаранне рэакцыйнай сумесі ці ланцуговы выбух. Значны ўклад у развіццё тэорыі Л.х.р. М.М.Сямёнава і С.Н.Хіншэлвуда. Гл. таксама Кінетыка хімічная.
Літ.:
Семенов Н.Н. О некоторых проблемах химической кинетики и реакционной способности. 2 изд. М., 1958;
Яго ж. Развитие теории цепных реакций и теплового воспламенения. М., 1969.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АПАНАСЕ́ВІЧ Павел Андрэевіч
(н. 14.7.1929, в. Стараселле Докшыцкага р-на Віцебскай вобл.),
бел. фізік. Акад. (1984, чл.-кар. 1980) АН Беларусі. Засл. дз. нав. Беларусі (1955). Д-рфіз.-матэм. н. (1974), праф. (1977). Скончыў БДУ (1954). З 1955 у Ін-це фізікі АН Беларусі (з 1987 дырэктар). Навук. працы па оптыцы і лазернай фізіцы. Развіў тэорыю ўздзеяння магутнага выпрамянення на спектральна-аптычныя характарыстыкі атамаў і малекул, устанавіў шэраг заканамернасцяў узаемадзеяння патокаў святла ў розных асяроддзях, генерацыі звышкароткіх светлавых імпульсаў, вымушанага камбінацыйнага рассеяння. Распрацаваў шэраг метадаў нелінейнай спектраскапіі і кіравання параметрамі лазерных патокаў. Дзярж. прэмія Беларусі 1978. Дзярж. прэмія СССР 1982.
Тв.:
Таблицы распределения энергии и фотонов в спектре равновесного излучения. Мн., 1961 (разам з В.С.Айзенштатам);
Основы теории взаимодействия света с веществами. Мн., 1977.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЕНЕТЫ́ЧНАЯ ІНФАРМА́ЦЫЯ,
праграма развіцця арганізма, атрыманая ад продкаў і закладзеная ў спадчынных структурах — генах. Запісана паслядоўнасцю нуклеатыдаў малекул нуклеінавых к-т (ДНК, у некат. вірусаў таксама РНК). Мае звесткі пра будову ўсіх ферментаў, структурных бялкоў і РНК клеткі, а таксама пра рэгуляцыю іх сінтэзу. Генетычная інфармацыя, якая счытваецца ў працэсе трансляцыі, складаецца са значэнняў трыплетаў генетычнага кода і ўключае знакі пачатку і заканчэння бялковага сінтэзу. У шматклетачных арганізмаў пры палавым размнажэнні генетычная інфармацыя перадаецца з пакалення ў пакаленне праз палавыя клеткі, у пракарыятычных мікраарганізмаў — праз трансдукцыю і трансфармацыю. Адрозніваюць 3 тыпы працэсаў пераносу генетычнай інфармацыі: агульны, уласцівы любым клеткам арганізма; спецыялізаваны (напр., у клетках, пашкоджаных вірусамі, генет. матэрыял якіх складаецца з РНК); забаронены перанос — працэсы, якія раней ніколі не былі зарэгістраваны (ад бялку да ДНК і РНК; ад бялку да бялку).
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
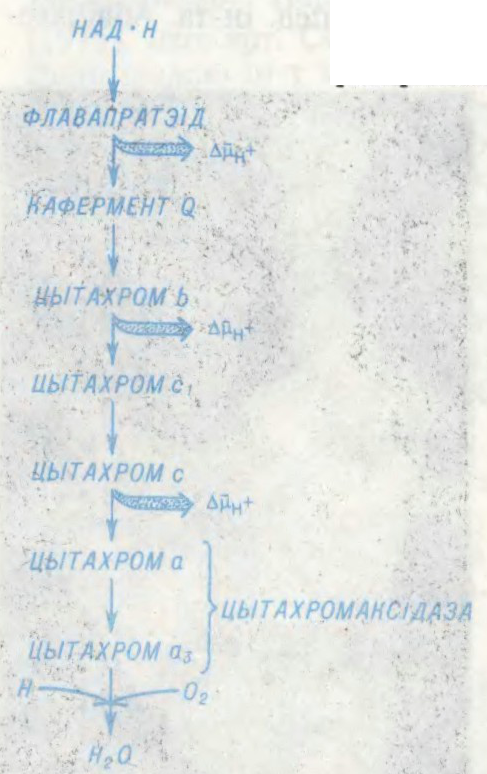
АКІСЛЯ́ЛЬНАЕ ФАСФАРЫЛІ́РАВАННЕ,
біясінтэз малекул адэназінтрыфосфарнай к-ты (АТФ) з адэназіндыфосфарнай (АДФ) і фосфарнай к-т за кошт энергіі акіслення субстрату: АДФ + H3PO4 → АТФ + H2O (гл.Акісленне біялагічнае). Асн. субстратам для акісляльнага фасфарыліравання служаць малекулы арган. рэчываў, пераважна арган.к-т, якія ўтвараюцца ў трыкарбонавых кіслот цыкле. Акісляльнае фасфарыліраванне адкрыта сав. біяхімікам У.А.Энгельгартам (1930). Адбываецца ў жывых клетках (у мітахондрыях) пры пераносе электронаў і пратонаў па дыхальным ланцугу з удзелам значнай колькасці дыхальных ферментаў і каферментаў (ферментны комплекс АТФ-сінтэтаза). Паказчык эфектыўнасці акісляльнага фасфарыліравання — адносіны колькасці фасфату, звязанага АДФ пры акісляльным фасфарыліраванні, да паглынутага кіслароду (P/O). Малекула АТФ утвараецца пры пераносе 2 электронаў праз пункт энергет. злучэння, у якім адбываецца утылізацыя энергіі, што вызваляецца. У выніку акісляльнага фасфарыліравання ў клетках назапашваецца АТФ — важнейшае макраэргічнае злучэнне, якое забяспечвае розныя працэсы жыццядзейнасці.