ВЫВЕ́ТРЫВАННЕ,
працэс
У выніку
На Беларусі ў сучасных
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ВЫВЕ́ТРЫВАННЕ,
працэс
У выніку
На Беларусі ў сучасных
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КАМЕ́ННЫ ВУ́ГАЛЬ,
цвёрды гаручы карысны выкапень расліннага паходжання, разнавіднасць вуглёў выкапнёвых, прамежкавая паміж бурым вугалем і антрацытам. Мае ў гаручай масе 75—92% вугляроду, 2,5—5,7 вадароду, 1,5—15% кіслароду,
Важнай характарыстыкай К.в. з’яўляецца ступень яго рэгіянальнага метамарфізму, або узровень пераўтварэння
У.Я.Бардон.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АКІСЛЕ́ННЕ БІЯЛАГІ́ЧНАЕ,
біяхімічны працэс, сукупнасць акісляльна-аднаўляльных рэакцый. Адбываецца ва ўсіх жывых клетках (пераважна ў мітахондрыях), складае аснову тканкавага дыхання і браджэння.
Вывучэнне працэсаў акіслення ў арганізме пачалося ў 18
На Беларусі розныя аспекты акіслення біялагічнага вывучаюць у ін-тах біяарганічнай хіміі, фотабіялогіі, фізіялогіі, біяхіміі (Гродна)
Літ.:
Кривобокова С.С. Биологическое окисление: Ист. очерк
Ленинджер А. Основы биохимии:
Строев Е.А. Биологическая химия.
Березов Т.Т., Коровкин Б.Ф. Биологическая химия. 2 изд.
М.М.Філімонаў.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АБМЕ́Н РЭ́ЧЫВАЎ,
сукупнасць
Ператварэнні рэчываў ад моманту іх паступлення ў арганізм да ўтварэння канчатковых прадуктаў распаду складаюць сутнасць
Літ.:
Ленинджер А. Основы биохимии:
Страйер Л. Биохимия:
Я.В.Малашэвіч.
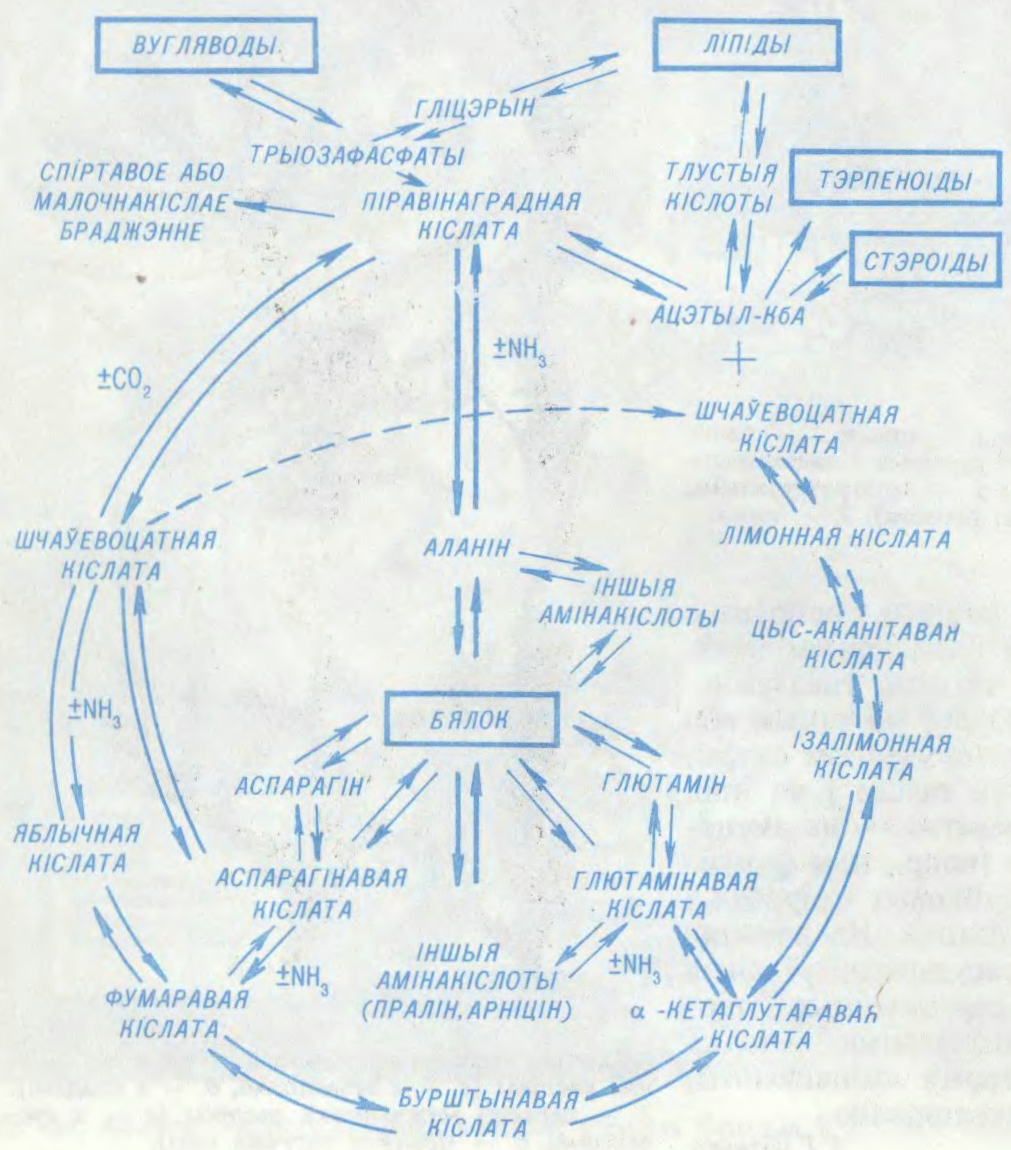
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГАРЭ́ЦКІ (Гаўрыла Іванавіч) (10.4.1900,
Тв.:
Народны прыбытак Беларусі.
Аллювий великих антропогеновых прарек Русской равнины. Прареки Камского бассейна.
Аллювиальная летопись великого Пра-Днепра.
Асноўныя вынікі вывучэння антрапагену Беларусі ў Акадэміі навук
Особенности палеопотамологии ледниковых областей (на примере Белорусского Понеманья).
Палеопотамологические эскизы Палео-Дона и Пра-Дона.
Літ.:
Бібліяграфія навуковых прац акадэміка
Т.В.Якубоўская.

Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЕАБАТА́НІКА (ад геа... + батаніка),
навука пра расліннае покрыва Зямлі, яго структурныя элементы — раслінныя згуртаванні (фітацэнозы); раздзел батанікі. Вывучае заканамернасці фарміравання, склад, будову, развіццё і класіфікацыю раслінных згуртаванняў, іх уплыў на асяроддзе. Развіваецца ў цеснай сувязі з комплексам
Як
На Беларусі геабат. работы праводзіліся з канца 19 —
Літ.:
Юркевич И.Д., Гельтман В.С. География, типология и районирование лесной растительности Белоруссии.
Юркевич И.Д., Голод Д.С., Адерихо В.С. Растительность Белоруссии, ее картографирование, охрана и использование.
Дз.С.Голад.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БЯЛКО́ВЫ АБМЕ́Н,
сукупнасць
Пры
Літ.:
Строев Е.А. Биологическая химия.
Николаев А.Я. Биологическая химия.
Березов Т.Т., Коровкин Б.Ф. Биологическая химия. 2 изд.
В.К.Кухта.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
КУ́КА АСТРАВЫ́ (Cook Islands). У цэнтральнай
Заселены палінезійцамі ў 1-м
Літ.:
Малаховский К.В. История островов Кука.
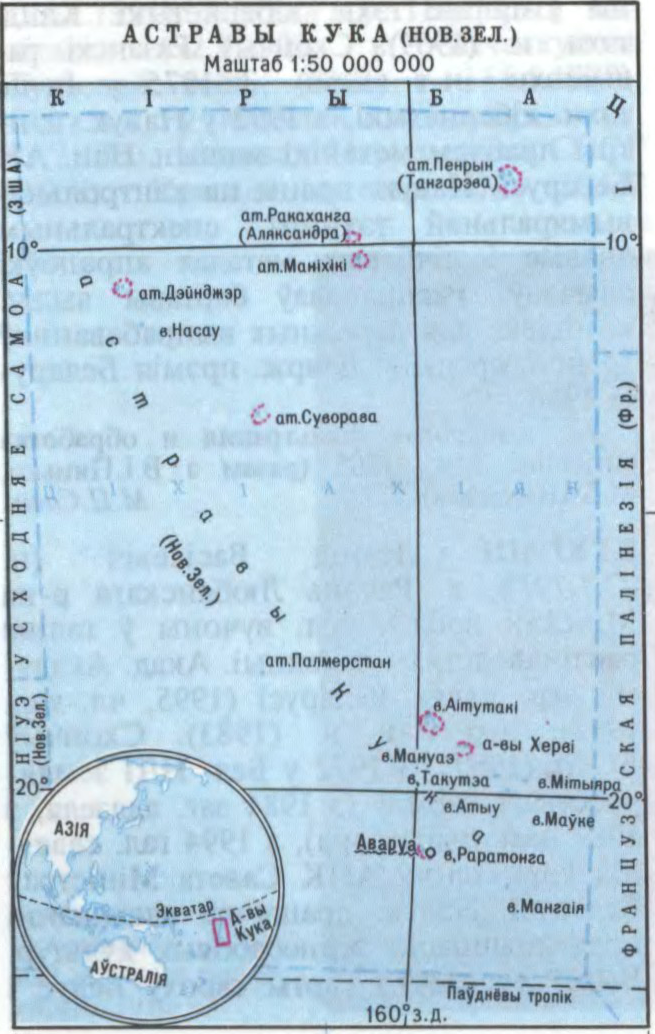
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АНАТО́МІЯ (ад
навука пра форму і будову асобных органаў, сістэм і арганізма ў цэлым; раздзел марфалогіі. Адрозніваюць анатомію чалавека (антрапатомію), анатомію жывёл (заатомію) і анатомію
У сваім развіцці
На Беларусі 1-е анатаміраванне цела праведзена ў 1586 у Гродне для ўдакладнення прычыны смерці караля Стафана Баторыя. Станаўленне сучаснай
Літ.:
Анатомия человека.
Акаевский А.И. Анатомия домашних животных. 3 изд.
П.І.Лабко.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АСМАТЫ́ЧНЫ ЦІСК,
лішкавы гідрастатычны ціск раствору, які перашкаджае дыфузіі растваральніку праз паўпранікальную перагародку; тэрмадынамічны параметр. Характарызуе імкненне раствору да зніжэння канцэнтрацыі пры сутыкненні з чыстым растваральнікам пры сустрэчнай дыфузіі малекул растворанага рэчыва і растваральніку. Абумоўлены змяншэннем хімічнага патэнцыялу растваральніку ў прысутнасці растворанага рэчыва. Роўны лішкаваму вонкаваму ціску, які неабходна прыкласці з боку раствору, каб спыніць осмас. Вымяраецца ў паскалях.
Вымярэнні асматычнага ціску пачаў у 1877
Асматычны ціск адыгрывае важную ролю ў жыццядзейнасці жывых клетак і арганізмаў. У клетках і
Літ.:
Курс физической химии.
Пасынский А.Г. Коллоидная химия. 3 изд.
Гриффин Д., Новик
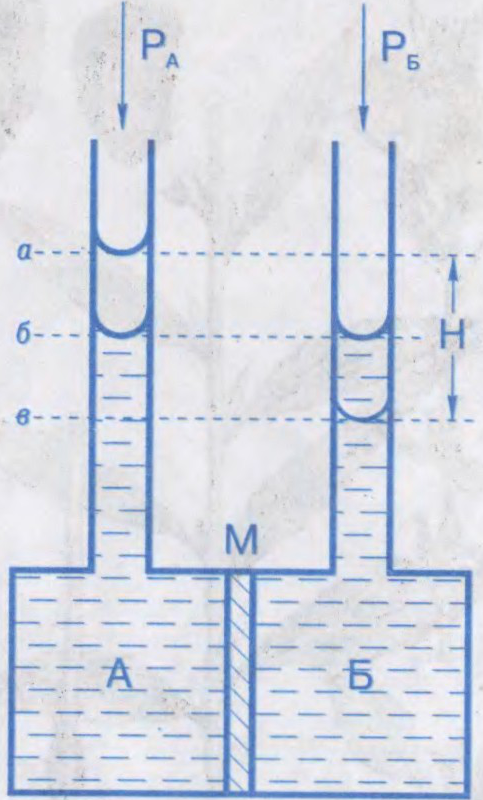
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)