свячэнне жывых арганізмаў, абумоўленае біяхім. працэсамі; від хемалюмінесцэнцыі. Уласціва некаторым бактэрыям, ніжэйшым раслінам, грыбам, насякомым (жукі-светлякі і інш.), беспазваночным, рыбам. Вельмі пашырана сярод марскіх жывёл.
У большасці выпадкаў біялюмінісцэнцыя ўзнікае ў выніку ферментатыўнага акіслення асобных рэчываў — люцыферынаў. Частка малекул люцыферынаў за кошт вызваленай пры гэтым хім. энергіі пераходзіць ва ўзбуджаны стан, пры вяртанні ў асн. стан яны выпраменьваюць святло. Біялюмінісцэнцыя выкарыстоўваецца для асвятлення і прынады здабычы (напр., у глыбакаводных рыб), для перасцярогі, адпужвання або адцягвання ўвагі драпежнікаў, у якасці сігналу для сустрэчы самцоў і самак у шлюбны перыяд (розныя віды светлякоў).
Літ.:
Мак-Элрой У.Д., Зелигер Г.Г. Происхождение и развитие биолюминесценции // Горизонты биохимии: Пер. с англ. М., 1964;
Тарусов Б.Н. Сверхслабое свечение живых организмов. М., 1972.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГІПЕРГУ́К,
пругкія хвалі з частатой 109—1013Гц. Па фізічнай прыродзе не адрозніваецца ад ультрагуку (2·104—109Гц). Існуе гіпергук прыродны (цеплавыя ваганні крышталічнай рашоткі) і штучны (генерыруецца пры дапамозе спец. выпрамяняльнікаў; гл.П’езаэлектрычнасць, Магнітастрыкцыя).
Пругкія хвалі распаўсюджваюцца ў асяроддзі, калі іх даўжыні большыя за даўжыню свабоднага прабегу малекул у газах ці міжатамных адлегласцей у вадкіх і цвёрдых целах. Таму ў газах, у т. л. ў паветры, пры нармальных умовах гіпергук не распаўсюджваецца, у вадкасцях хутка затухае; параўнальна добрыя праваднікі гіпергуку — монакрышталі пры нізкіх т-рах. Гіпергук выкарыстоўваюць для даследавання стану рэчыва, асабліва ў фізіцы цвёрдага цела, для стварэння акустычных ліній затрымкі ў ЗВЧ дыяпазоне і інш. прылад акустаэлектронікі і акустаоптыкі.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ЛАНЦУГО́ВЫЯ ХІМІ́ЧНЫЯ РЭА́КЦЫІ,
хімічныя рэакцыі, у якіх узнікненне прамежкавай актыўнай часціцы (атама, свабоднага радыкала, іона) і яе рэгенерацыя (узнаўленне) у кожным элементарным акце рэакцыі выклікаюць вялікую колькасць (ланцуг) пераўтварэнняў зыходных рэчываў у прадукты рэакцыі. Тыповыя Л.х.р. — тэрмічны крэкінг, піроліз, акісленне, радыкальная полімерызацыя, галагеніраванне.
Л.х.р. складаецца з некалькіх элементарных стадый: зараджэнне ланцуга (ініцыіраванне) — утварэнне актыўных часціц (АЧ), якое адбываецца, напр., у выніку дысацыяцыімалекул; працяг ланцуга — узаемадзеянне АЧ з зыходнымі рэчывамі, якое прыводзіць да ўтварэння малекул прадукту рэакцыі і новых АЧ; абрыў ланцуга («знікненне» АЧ) у выніку рэкамбінацыі свабодных радыкалаў, узаемадзеяння АЧ са сценкамі пасудзін ці з інгібітарамі. Адрозніваюць Л.х.р. неразгалінаваныя і разгалінаваныя. У неразгалінаваных на кожную АЧ, якая расходуецца пры працягу ланцуга, узнікае адна актыўная часціца, напр., рэакцыя хларыравання вадароду. Пры ініцыіраванні ўтвараецца свабодны атам хлору (Cl), які ўзаемадзейнічае з малекулай вадароду H2(Cl+H2 → HCl+H) з утварэннем малекулы хлорыстага вадароду HCl і свабоднага атама вадароду, які ўзаемадзейнічае з малекулай Cl2(H+Cl2 → HCl+Cl), і г.д. У разгалінаваных Л.х.р. на адну АЧ, расходаваную пры працягу ланцуга, узнікае некалькі АЧ (2 і больш), адна з іх працягвае першы ланцуг, а інш. пачынаюць новыя, што прыводзіць да разгалінавання. Калі скорасць разгалінавання меншая за скорасць знікнення АЧ, разгалінаваная Л.х.р. адбываецца ў стацыянарным рэжыме. Пры нестацыянарным рэжыме (скорасць знікнення АЧ меншая за скорасць разгалінавання) агульная скорасць ланцуговага працэсу імкліва ўзрастае. Пераход да нестацыянарнага рэжыму адбываецца скачком і разглядаецца як самазагаранне рэакцыйнай сумесі ці ланцуговы выбух. Значны ўклад у развіццё тэорыі Л.х.р. М.М.Сямёнава і С.Н.Хіншэлвуда. Гл. таксама Кінетыка хімічная.
Літ.:
Семенов Н.Н. О некоторых проблемах химической кинетики и реакционной способности. 2 изд. М., 1958;
Яго ж. Развитие теории цепных реакций и теплового воспламенения. М., 1969.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АПАНАСЕ́ВІЧ Павел Андрэевіч
(н. 14.7.1929, в. Стараселле Докшыцкага р-на Віцебскай вобл.),
бел. фізік. Акад. (1984, чл.-кар. 1980) АН Беларусі. Засл. дз. нав. Беларусі (1955). Д-рфіз.-матэм. н. (1974), праф. (1977). Скончыў БДУ (1954). З 1955 у Ін-це фізікі АН Беларусі (з 1987 дырэктар). Навук. працы па оптыцы і лазернай фізіцы. Развіў тэорыю ўздзеяння магутнага выпрамянення на спектральна-аптычныя характарыстыкі атамаў і малекул, устанавіў шэраг заканамернасцяў узаемадзеяння патокаў святла ў розных асяроддзях, генерацыі звышкароткіх светлавых імпульсаў, вымушанага камбінацыйнага рассеяння. Распрацаваў шэраг метадаў нелінейнай спектраскапіі і кіравання параметрамі лазерных патокаў. Дзярж. прэмія Беларусі 1978. Дзярж. прэмія СССР 1982.
Тв.:
Таблицы распределения энергии и фотонов в спектре равновесного излучения. Мн., 1961 (разам з В.С.Айзенштатам);
Основы теории взаимодействия света с веществами. Мн., 1977.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ГЕНЕТЫ́ЧНАЯ ІНФАРМА́ЦЫЯ,
праграма развіцця арганізма, атрыманая ад продкаў і закладзеная ў спадчынных структурах — генах. Запісана паслядоўнасцю нуклеатыдаў малекул нуклеінавых к-т (ДНК, у некат. вірусаў таксама РНК). Мае звесткі пра будову ўсіх ферментаў, структурных бялкоў і РНК клеткі, а таксама пра рэгуляцыю іх сінтэзу. Генетычная інфармацыя, якая счытваецца ў працэсе трансляцыі, складаецца са значэнняў трыплетаў генетычнага кода і ўключае знакі пачатку і заканчэння бялковага сінтэзу. У шматклетачных арганізмаў пры палавым размнажэнні генетычная інфармацыя перадаецца з пакалення ў пакаленне праз палавыя клеткі, у пракарыятычных мікраарганізмаў — праз трансдукцыю і трансфармацыю. Адрозніваюць 3 тыпы працэсаў пераносу генетычнай інфармацыі: агульны, уласцівы любым клеткам арганізма; спецыялізаваны (напр., у клетках, пашкоджаных вірусамі, генет. матэрыял якіх складаецца з РНК); забаронены перанос — працэсы, якія раней ніколі не былі зарэгістраваны (ад бялку да ДНК і РНК; ад бялку да бялку).
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
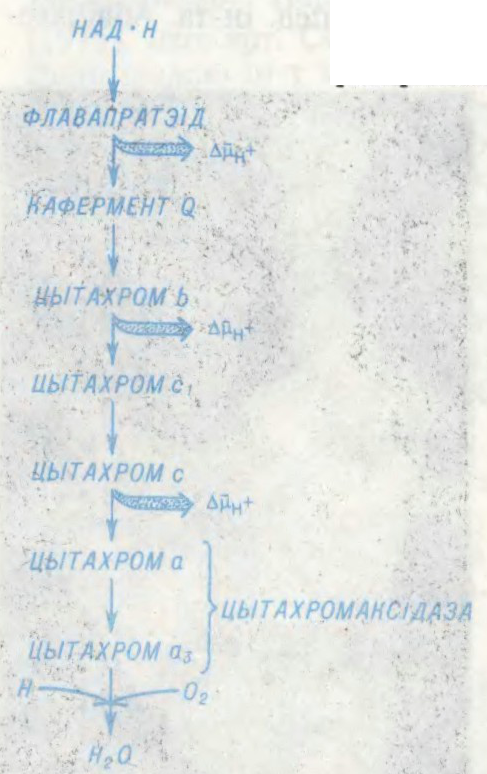
АКІСЛЯ́ЛЬНАЕ ФАСФАРЫЛІ́РАВАННЕ,
біясінтэз малекул адэназінтрыфосфарнай к-ты (АТФ) з адэназіндыфосфарнай (АДФ) і фосфарнай к-т за кошт энергіі акіслення субстрату: АДФ + H3PO4 → АТФ + H2O (гл.Акісленне біялагічнае). Асн. субстратам для акісляльнага фасфарыліравання служаць малекулы арган. рэчываў, пераважна арган.к-т, якія ўтвараюцца ў трыкарбонавых кіслот цыкле. Акісляльнае фасфарыліраванне адкрыта сав. біяхімікам У.А.Энгельгартам (1930). Адбываецца ў жывых клетках (у мітахондрыях) пры пераносе электронаў і пратонаў па дыхальным ланцугу з удзелам значнай колькасці дыхальных ферментаў і каферментаў (ферментны комплекс АТФ-сінтэтаза). Паказчык эфектыўнасці акісляльнага фасфарыліравання — адносіны колькасці фасфату, звязанага АДФ пры акісляльным фасфарыліраванні, да паглынутага кіслароду (P/O). Малекула АТФ утвараецца пры пераносе 2 электронаў праз пункт энергет. злучэння, у якім адбываецца утылізацыя энергіі, што вызваляецца. У выніку акісляльнага фасфарыліравання ў клетках назапашваецца АТФ — важнейшае макраэргічнае злучэнне, якое забяспечвае розныя працэсы жыццядзейнасці.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
АДБО́РУ ПРА́ВІЛЫў фізіцы,
умовы, што вызначаюць магчымасць пераходу квантавых сістэм (ядраў, атамаў, малекул і інш.) з пачатковага стану ў канчатковы пры фіз. працэсах, звязаных з выпрамяненнем і паглынаннем энергіі.
Адбору правілы выражаюць выкананне пэўных захавання законаў у дадзеным працэсе і фармулююцца ў выглядзе суадносін паміж квантавымі лікамі. Аснова тэарэт. вызначэння адбору правілаў — патрабаванне адрознення ад нуля імавернасці пераходу паміж пач. і канчатковым станамі сістэмы, напр., імавернасць дыпольных пераходаў, звязаных з выпрамяненнем святла атамам, адрозніваецца ад нуля пры змене квантавых лікаў; ΔL = ±1, Δs = 0, ΔI = 0 або ±1 (за выключэннем, калі I = 0 у пач. і канчатковым станах), дзе I, L і s — адпаведна квантавыя лікі поўнага моманту імпульсу электроннай абалонкі, арбітальнага моманту і агульнага спінавага моманту электронаў. Пераходы, якія падпарадкоўваюцца адбору правілам дыпольнага выпрамянення, наз. дазволенымі, у адваротным выпадку — забароненымі (іх імавернасць у атамах вельмі малая). Адпаведныя адбору правілы існуюць у ядз. спектраскапіі і фізіцы элементарных часціц.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БІЯЛАГІ́ЧНЫЯ МЕМБРА́НЫ,
тонкія пагранічныя паўпранікальныя бялкова-ліпідныя малекулярныя структуры на паверхні клетак і субклетачных часціц (арганел), а таксама канальцаў і пузыркоў, што пранізваюць пратаплазму. Таўшчыня не больш за 10 нм. Складзены з бімалекулярнага слоя ліпідаў (пераважна фосфаліпідаў), у якім размешчаны розныя мембранныя бялкі, гетэрагенныя макрамалекулы (глікапратэіды, глікаліпіды) і розныя мінорныя кампаненты (нуклеінавыя к-ты, каферменты, караціноіды і інш.), што адказваюць за б.ч. мембранных функцый. У залежнасці ад віду біялагічныя мембраны выконваюць разнастайныя функцыі: актыўны транспарт іонаў і розных рэчываў (соляў, цукроў, амінакіслот і інш. прадуктаў метабалізму) і яго рэгуляванне: агульная і выбарчая дыфузія невял. малекул і іонаў (усе віды біялагічных мембран); электраізаляванне (біялагічныя мембраны міэліну); генерацыя нерв. імпульсу (біялагічныя мембраны нерв. клетак); пераўтварэнне светлавой энергіі ў хім. энергію АТФ — адэназінтрыфосфарнай кіслаты (біялагічныя мембраны хларапластаў); пераўтварэнне энергіі біял. акіслення ў хім. энергію макраэргічных фасфатных сувязяў у малекуле АТФ (біялагічныя мембраны мітахондрый); фагацытоз, пінацытоз; антыгенныя рэакцыі (біялагічныя мембраны спецыялізаваных клетак); бар’ерная, каталітычная функцыі і інш. Падтрымліваючы нераўнамернасць размеркавання іонаў калію, натрыю, хлору і інш. паміж пратаплазмай і навакольным асяроддзем, біялагічныя мембраны садзейнічаюць узнікненню рознасці біяэлектрычных патэнцыялаў. Гл. таксама Клетачныя мембраны.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
БІЯРЫТМАЛО́ГІЯ
(ад бія... + рытм + ...логія),
хранабіялогія, раздзел біялогіі, які вывучае цыклічныя працэсы ў біял. сістэмах. Як навука ўзнікла ў 2-й пал. 20 ст. У 1950—60-я г. сфарміраваліся ўяўленні аб фізіял. рытмах — высокачастотных перыядычных працэсах эндагеннага паходжання, што рэалізуюцца пераважна на біяхім., клетачным і тканкавым узроўнях арганізацыі (метабалічныя цыклы, рытмы сардэчны, дыхання, электрапатэнцыялаў мозга), і біялагічных рытмах, перыядычнасць якіх задаецца рытмам вонкавых сінхранізавальных фактараў, але з рэалізацыяй на ўсіх узроўнях (ад малекул да біясферы). Спалучэнне «эндагеннасці» і «экзагеннасці» фарміравання біярытмаў — адна з асн. праблем біярытмалогіі. Існуе агульная тэндэнцыя да павелічэння даўжыні перыядаў біярытмаў па меры таго, як ускладняліся біял. сістэмы. З біярытмалогіі вылучыліся хранамарфалагічны кірунак у вывучэнні структурнай арганізацыі біял. сістэм, касм. хранабіялогія; на стыку біярытмалогіі і клінічнай медыцыны — хранамедыцына, якая вывучае ўзаемасувязі біярытмаў з цячэннем розных хвароб і інш.мед. аспекты.
Беларуская Энцыклапедыя (1996—2004, правапіс да 2008 г., часткова)
ВУГЛЯВО́ДНЫ АБМЕ́Н,
сукупнасць хім. працэсаў дэградацыі (катабалізму) і біясінтэзу (анабалізму) вугляводаў у арганізме. На 1-й стадыі катабалізму пры ўдзеле стрававальных ферментаў складаныя поліцукрыды і алігацукрыды распадаюцца да монацукрыдаў (гексоз і пентоз), якія на 2-й стадыі расшчапляюцца да аднаго і таго ж трохвугляроднага прамежкавага прадукту — пірувату (гліколіз), а потым у аэробных умовах да двухвугляроднай формы — ацэтыльнай групы ацэтылкаферменту A (гл.Трыкарбонавых кіслот цыкл). У анаэробных умовах піруват у большай частцы клетак жывёльных і раслінных тканак аднаўляецца да лактату, а ў клетках дражджэй у ходзе спіртавога браджэння ператвараецца ў этылавы спірт і вуглякіслы газ. На 3-й стадыі ацэтыльная група ацэтылкаферменту A уступае ў цыкл лімоннай к-ты — агульны канчатковы шлях, на якім усе віды малекул вугляводаў акісляюцца да вуглякіслага газу. Дэградацыя вугляводаў у арганізме суправаджаецца вызваленнем значнай энергіі, якая расходуецца на розныя працэсы жыццядзейнасці. Біясінтэз вугляводаў у жывых клетках можа адбывацца шляхам глюканеагенезу (сінтэз глюкозы ў клетках печані, які ўключае 9 з 11 ферментацыйных рэакцый, што ўдзельнічаюць у яе раскладзе) і шляхам ператварэння простых вугляводаў у больш складаныя аліга- і поліцукрыды.